0597
Diffusivity and kurtosis time-dependence changes in the rat brain during somatosensory evoked response1Dept. of Radiology, Lausanne University Hospital (CHUV) and University of Lausanne, Lausanne, Switzerland
Synopsis
Keywords: Microstructure, Gray Matter, Microstructure, Permeability, fMRI (task based), Multimodal, Small Animals, Preclinical
Motivation: The microstructure-function relationship is essential for our understanding of the healthy and pathological human brain.
Goal(s): In this context, the goal of our study was to investigate microstructural changes arising in the brain during neuronal activity.
Approach: Differences in mean diffusivity MD and mean kurtosis MK time-dependence between rest and active states were assessed in the somatosensory cortex following rat forepaw stimulation.
Results: While no changes were found in control cortical regions, the BOLD-fMRI positive cluster presented a significant decrease in MD and MK during activation. Interestingly, subcortical somatosensory relays displayed the opposite trend which could result from changes in inhibitory/excitatory balance.
Impact: Neuronal activity is accompanied by a myriad of microstructural changes. The diffusion-weighted signal sensitivity to underlying brain microstructure brings new perspectives into the structure-function relationship along with the promise of a functional contrast unbound from the current limitations of BOLD-fMRI.
Introduction
In diffusion MRI (dMRI), universal classes of tissue disorder1 allow the exploration of the time-dependent nature of diffusion in tissues and offer valuable insights into microstructural properties and dynamic phenomena2–6. Neuronal activity is accompanied by several physiological and biochemical processes, which lead to cellular swelling7–10 and increased transmembrane permeability11. In this study we propose to investigate differences in mean diffusivity MD(∆) and mean kurtosis MK(∆) that arise during neuronal activation on account of morphological and metabolic changes. Forepaw electrical stimulation is used in adult rats to generate a somatosensory response. BOLD-fMRI is acquired as a control for the activation and the BOLD activation cluster is employed to define the region-of-interest (ROI) in which main diffusion changes between the rest and active states are assessed. Other ROIs in the brain devoid of BOLD response are also examined.Methods
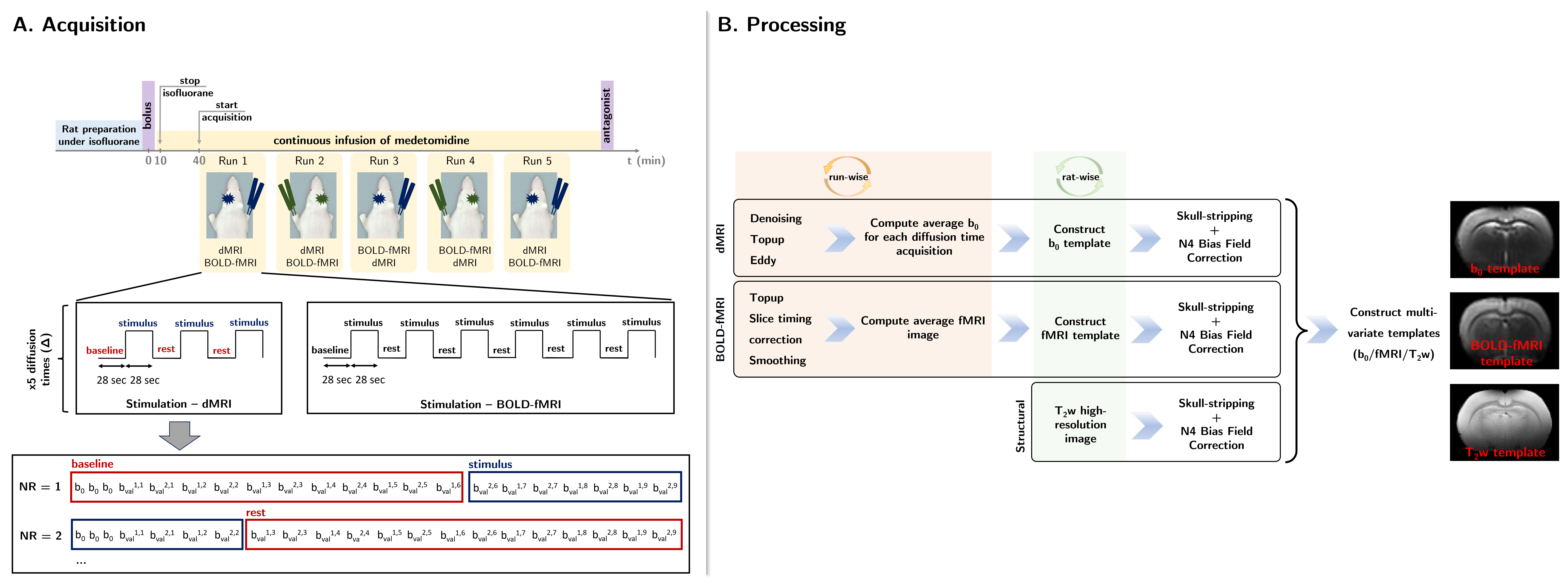
Animal preparation: Experiments were performed in vivo on six female rats (241±16g) on a 14.1T system, using a home-built surface transceiver RF-coil. Electrodes were implanted in the rat forepaws12 under isoflurane anesthesia. Anesthesia was gradually switched to medetomidine and the dMRI/BOLD-fMRI protocol started at fixed time-delay after medetomidine bolus (Fig.1A).Paradigm design: Stimulation consisted in 0.3ms pulses at 2mA and 9Hz frequency, triggered by the scanner. Each run consisted in blocks of 28s stimulation and 28s rest (Fig.1A). Left and right paw stimulation were alternated.
Structural: RARE-T2w sequences (TEeff/TR=6/2500ms, matrix size 160×160x45; in-plane resolution 125x125μm2; slice thickness 0.5mm) were used for anatomical reference.
dMRI: PGSE-EPI sequences (TE/TR=48/2000ms; matrix size 80x80x21; slice thickness 0.75µm; in-plane resolution 0.25x0.25µm2; δ=4ms) were used. For each run there were five dMRI acquisitions (Δ=9.5;15;20;25;30ms). Each acquisition included 21 measurements13 (3 b0, two b-values bvali,j, where i=1;2ms/µm2 and j=9 directions) repeated 4 times while alternating between rest and stimulation (Fig.1A). Data underwent denoising14, topup15 and eddy16 corrections (Fig.1B) before the estimation of axially-symmetric diffusion and kurtosis tensors17. MD(∆)&MK(∆) were evaluated using functional forms of disorder and exchange.
BOLD-fMRI: A gradient-echo EPI sequence was used (TE/TR=11.1/1000ms) with the same geometry as the dMRI acquisitions. Data underwent topup15, slice timing correction and spatial smoothing prior to analyzing the BOLD response in SPM12.
Template generation: ANTs18 was employed to generate multivariate b0/BOLD-fMRI/T2w templates (Fig.1B), which served as a common space for the propagation of ROIs from the Waxholm Space Atlas19, as well as the propagation of the BOLD cluster segmentations to the diffusion images.
Statistical analysis: A mixed effects model was used to determine the significance of differences in MD&MK across diffusion times and across state (rest vs. active). Subjects and run numbers were modeled as nested random effects.
Results and Discussion
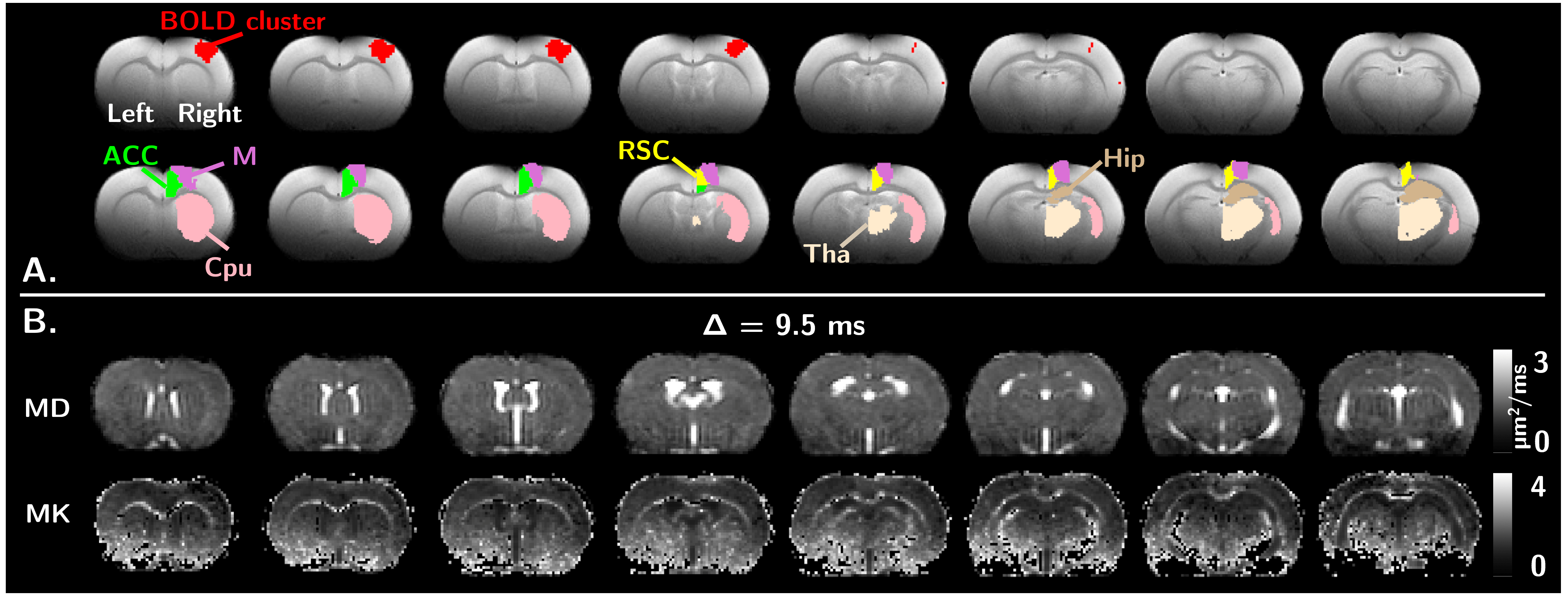
Only results for the right hemisphere are presented, where contralateral designates the left forepaw stimulation (BOLD response in the right somatosensory cortex - SC), and ipsilateral designates right forepaw stimulation (BOLD response in the left SC).Fig.2 shows an example of BOLD activation cluster and other atlas-defined ROIs investigated, along with examples of MD&MK maps.
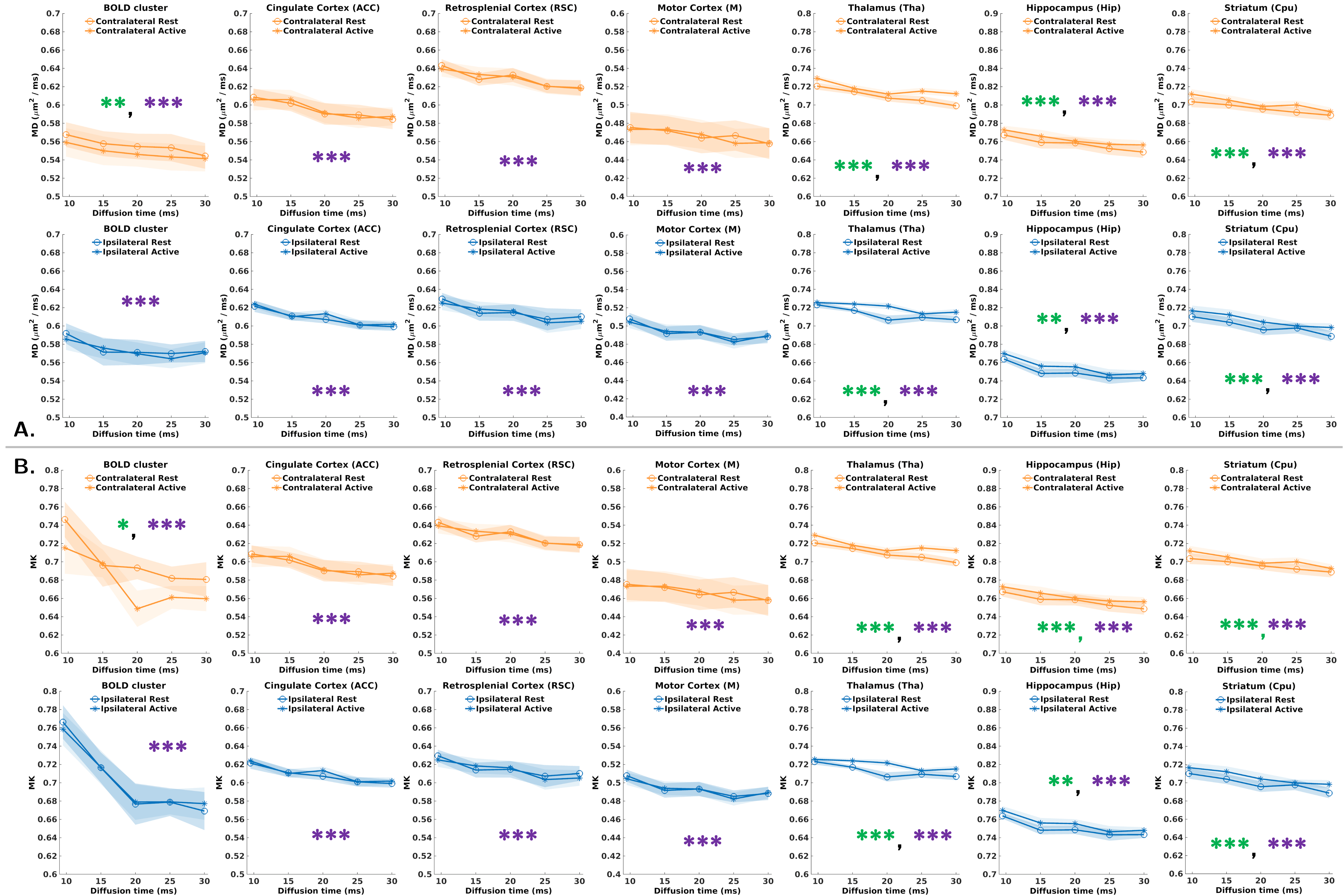
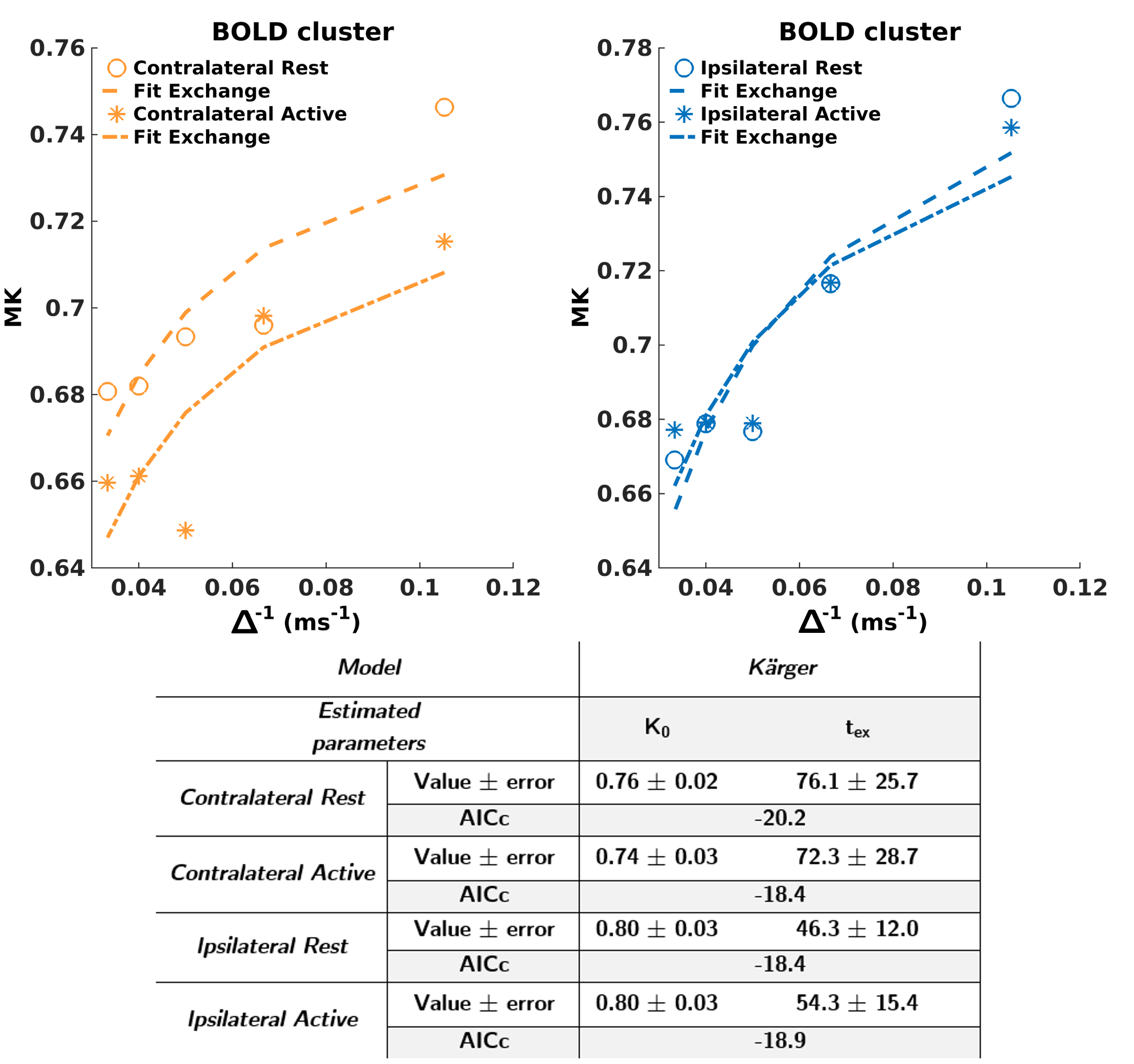
Fig.3 displays MD(∆)&MK(∆) in the BOLD-defined cluster and the other ROIs.
Rest/Stimulus differences: Statistically significant decrease in MD&MK in the active state was obtained in the contralateral BOLD cluster (green asterisks), while no differences were found ipsilaterally or in any cortical control region (cingulate, retrosplenial, motor). The decrease in MD during activation is in line with previous studies20,21 and attributed to cellular swelling7–10. MK is expected to result from competing mechanisms of increased restriction (MK increase) and permeability (MK decrease). The net decrease suggests permeability dominates.
Remarkably, a statistically significant increase in MD&MK was obtained in the thalamus, hippocampus, and striatum for contralateral and ipsilateral activations of SC, while these regions did not display significant BOLD response. The thalamus is responsible for relaying somatosensory signals22,23, with known temporal coherence between thalamus and SC during activation24, while the striatum25 and hippocampus26 are also part of the somatosensory relays. The increase in MD&MK during activation might reflect complex inhibitory and excitatory responses occurring in these brain regions during somatosensory input processing.
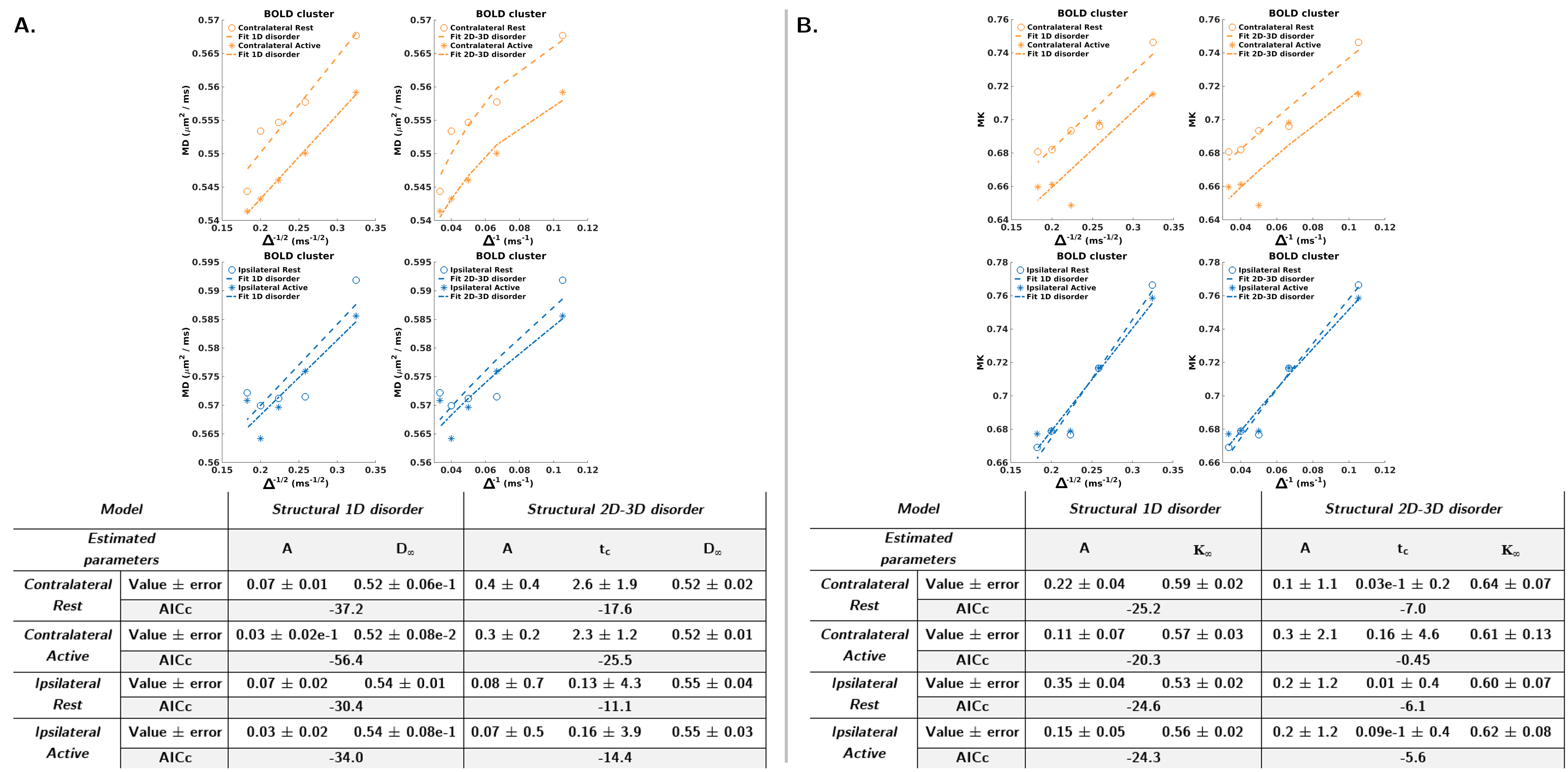
Measurable MD(∆)&MK(∆) was found in all ROIs and both conditions (Fig.3, purple asterisks). Lower AICc from the 1D-disorder model (Fig.4A) points towards a superior contribution from intracellular disorder to the MD temporal decay. For MK(∆), similar AICc for 1D-structural disorder (Fig.4B) and the Kärger (Fig.5) models outline the contribution of both effects, thus hampering a robust estimation of the exchange time from the Kärger model.
Conclusion
A significant decrease in MD(∆)&MK(∆) accompanies the activation of the somatosensory cortex and can be attributed to changes in both morphology and permeability. Remarkably, subcortical relays displayed opposite trends which could result from changes in inhibitory/excitatory balance.Acknowledgements
The authors thank Lucas Soustelle for sharing the diffusion sequence with the reversed phase encoding and for help with the implementation of the trigger in the diffusion sequence, Stefan Mitrea and Analina Hausin for their assistance with animal setup and monitoring, Bernard Lanz and Claudia Zanella for providing the RF coil. This work took place at the CIBM Center for Biomedical Imaging, a Swiss research center founded and supported by Lausanne University Hospital (CHUV), the University of Lausanne (UNIL), the Swiss Federal Institute of Technology (EPFL), the University of Geneva (UNIGE) and Geneva University Hospital (HUG) and was supported by the Swiss National Science Foundation under Eccellenza grant PCEFP2_194260.References
1. Novikov DS, Jensen JH, Helpern JA, Fieremans E. Revealing mesoscopic structural universality with diffusion. Proc Natl Acad Sci. 2014;111(14):5088-5093. doi:10.1073/pnas.1316944111
2. Fieremans E, Burcaw LM, Lee HH, Lemberskiy G, Veraart J, Novikov DS. In vivo observation and biophysical interpretation of time-dependent diffusion in human white matter. NeuroImage. 2016;129:414-427. doi:10.1016/j.neuroimage.2016.01.018
3. Reynaud O.
Time-Dependent Diffusion MRI in Cancer: Tissue Modeling and Applications. Front
Phys. 2017;5. https://www.frontiersin.org/articles/10.3389/fphy.2017.00058.
4. Lee HH, Papaioannou A, Novikov DS, Fieremans E. In vivo observation and biophysical interpretation of time-dependent diffusion in human cortical gray matter. NeuroImage. 2020;222:117054. doi:10.1016/j.neuroimage.2020.117054
5. Olesen JL, Østergaard L, Shemesh N, Jespersen SN. Diffusion time dependence, power-law scaling, and exchange in gray matter. Neuroimage. 2022;251:118976. doi:10.1016/j.neuroimage.2022.118976
6. Jelescu IO, de Skowronski A, Geffroy F, Palombo M, Novikov DS. Neurite Exchange Imaging (NEXI): A minimal model of diffusion in gray matter with inter-compartment water exchange. NeuroImage. 2022;256:119277. doi:10.1016/j.neuroimage.2022.119277
7. Saly V, Andrew RD. CA3 neuron excitation and epileptiform discharge are sensitive to osmolality. J Neurophysiol. 1993;69(6):2200-2208. doi:10.1152/jn.1993.69.6.2200
8. Andrew RD, Macvicar BA. Imaging cell volume changes and neuronal excitation in the hippocampal slice. Neuroscience. 1994;62(2):371-383. doi:10.1016/0306-4522(94)90372-7
9. Sun JY, Wu LG. Fast Kinetics of Exocytosis Revealed by Simultaneous Measurements of Presynaptic Capacitance and Postsynaptic Currents at a Central Synapse. Neuron. 2001;30(1):171-182. doi:10.1016/S0896-6273(01)00271-9
10. Tasaki I, Byrne PM. Rapid structural chances in nerve fibers evoked by electric current pulses. Biochem Biophys Res Commun. 1992;188(2):559-564. doi:10.1016/0006-291X(92)91092-5
11. Bai R, Springer CS, Plenz D, Basser PJ. Brain active transmembrane water cycling measured by MR is associated with neuronal activity. Magn Reson Med. 2019;81(2):1280-1295. doi:10.1002/mrm.27473
12. Reynaud O, da Silva AR, Gruetter R, Jelescu IO. Multi-slice passband bSSFP for human and rodent fMRI at ultra-high field. J Magn Reson. 2019;305:31-40. doi:10.1016/j.jmr.2019.05.010
13. Hansen B, Lund TE, Sangill R, Stubbe E, Finsterbusch J, Jespersen SN. Experimental considerations for fast kurtosis imaging. Magn Reson Med. 2016;76(5):1455-1468. doi:10.1002/mrm.26055
14. Veraart J, Fieremans E, Novikov DS. Diffusion MRI noise mapping using random matrix theory. Magn Reson Med. 2016;76(5):1582-1593. doi:10.1002/mrm.26059
15. Andersson JLR, Skare S, Ashburner J. How to correct susceptibility distortions in spin-echo echo-planar images: application to diffusion tensor imaging. NeuroImage. 2003;20(2):870-888. doi:10.1016/S1053-8119(03)00336-7
16. Andersson JLR, Sotiropoulos SN. An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage. 2016;125:1063-1078. doi:10.1016/j.neuroimage.2015.10.019
17. Hansen B, Lund TE, Sangill R, Jespersen SN. Experimentally and computationally fast method for estimation of a mean kurtosis. Magn Reson Med. 2013;69(6):1754-1760. doi:10.1002/mrm.24743
18. Avants BB, Tustison NJ, Song G, Cook PA, Klein A, Gee JC. A Reproducible Evaluation of ANTs Similarity Metric Performance in Brain Image Registration. NeuroImage. 2011;54(3):2033-2044. doi:10.1016/j.neuroimage.2010.09.025
19. Papp EA, Leergaard TB, Calabrese E, Johnson GA, Bjaalie JG. Waxholm Space atlas of the Sprague Dawley rat brain. NeuroImage. 2014;97:374-386. doi:10.1016/j.neuroimage.2014.04.001
20. Darquié A, Poline JB, Poupon C, Saint-Jalmes H, Le Bihan D. Transient decrease in water diffusion observed in human occipital cortex during visual stimulation. Proc Natl Acad Sci. 2001;98(16):9391-9395. doi:10.1073/pnas.151125698
21. Le Bihan D, Urayama S ichi, Aso T, Hanakawa T, Fukuyama H. Direct and fast detection of neuronal activation in the human brain with diffusion MRI. Proc Natl Acad Sci. 2006;103(21):8263-8268. doi:10.1073/pnas.0600644103
22. O’Reilly C, Iavarone E, Yi J, Hill SL. Rodent somatosensory thalamocortical circuitry: Neurons, synapses, and connectivity. Neurosci Biobehav Rev. 2021;126:213-235. doi:10.1016/j.neubiorev.2021.03.015
23. Sanganahalli BG, Thompson GJ, Parent M, et al. Thalamic activations in rat brain by fMRI during tactile (forepaw, whisker) and non-tactile (visual, olfactory) sensory stimulations. PLoS ONE. 2022;17(5):e0267916. doi:10.1371/journal.pone.0267916
24. Nunes D, Ianus A, Shemesh N. Layer-specific connectivity revealed by diffusion-weighted functional MRI in the rat thalamocortical pathway. NeuroImage. 2019;184:646-657. doi:10.1016/j.neuroimage.2018.09.050
25. Ramanathan S, Hanley JJ, Deniau JM, Bolam JP. Synaptic Convergence of Motor and Somatosensory Cortical Afferents onto GABAergic Interneurons in the Rat Striatum. J Neurosci. 2002;22(18):8158-8169. doi:10.1523/JNEUROSCI.22-18-08158.2002
26. Bellistri E, Aguilar J, Brotons-Mas JR, Foffani G, de la Prida LM. Basic properties of somatosensory-evoked responses in the dorsal hippocampus of the rat. J Physiol. 2013;591(Pt 10):2667-2686. doi:10.1113/jphysiol.2013.251892
Figures