0531
Biphasic training for awake imaging using a dual-imaging system reveals neurovascular uncoupling and anesthesia effects in healthy mice1Radiology and Biomedical Imaging, Yale University, New Haven, CT, United States, 2McGill, Montreal, QC, Canada
Synopsis
Keywords: Small Animals, Brain Connectivity, awake rodent imaging
Motivation: Most of rodent-fMRI is acquired under anesthesia, to minimize motion and stress. However, anesthesia hinders mouse-to-human translatability, since most of human fMRI is conducted whilst awake.
Goal(s): Here we aim to develop a biphasic protocol for conducting awake rodent-fMRI in mice and simultaneously recording mesoscopic calcium imaging data.
Approach: The animals undergo a first training and a refresher a few weeks later. Brain function is analyzed in simultaneous fMRI and mesoscopic calcium imaging measurements.
Results: Having a refresher training improves motion in the scanner. The two measures of brain function show interesting patterns over time, with partial agreement and some clear disagreement.
Impact: Rodent-fMRI is typically done under anesthesia to minimize stress and motion. This limits mouse-human translatability (since humans are usually scanned awake). Here, we develop a biphasic approach to train mice to awake imaging, using simultaneous fMRI and Calcium imaging measures.
Introduction
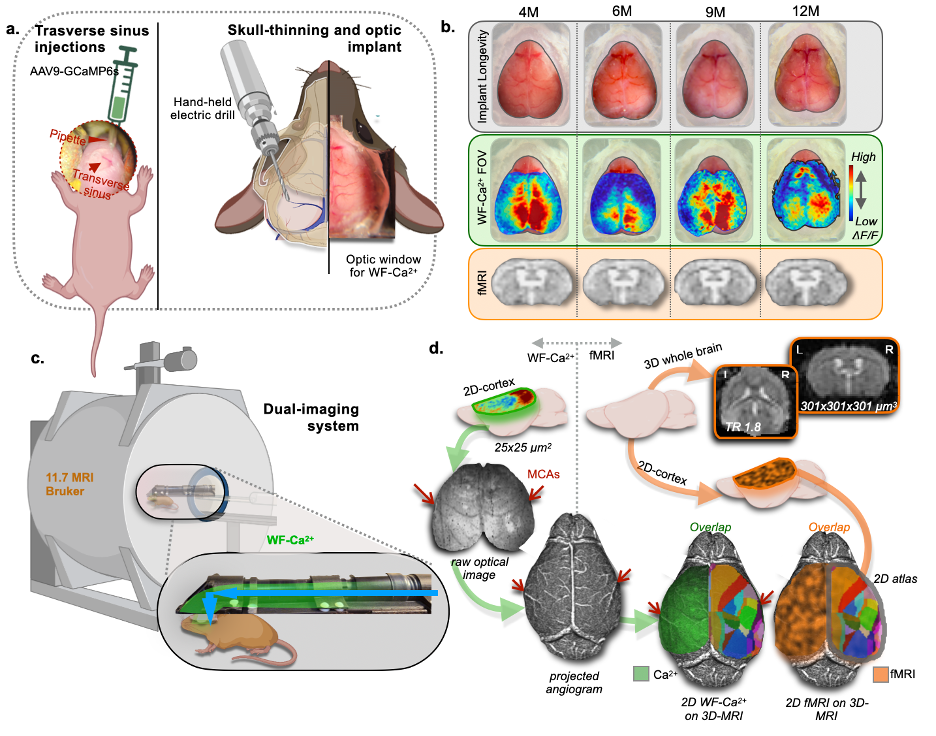
Awake imaging is gaining prominence in rodent fMRI, as it eliminates the confounding effects of anesthesia and improves the translatability of findings from mice to humans1-4. While fields like optical imaging routinely study awake mice, rodent fMRI is still mostly conducted on anesthetized mice, to minimize motion and stress. Here, we create a biphasic training approach to acclimate mice to awake imaging and demonstrate its feasibility using our multimodal dual-imaging system 5-7 (Fig. 1) with simultaneously acquired wide-field Ca2+ (WF-Ca2+) and fMRI data. We present a longitudinal design with four imaging sessions: the first two conducted under isoflurane anesthesia and the latter two involving training for awake imaging. Functional connectivity (FC) analyses are performed in both modalities and brain states, to explore neurovascular coupling.Methods
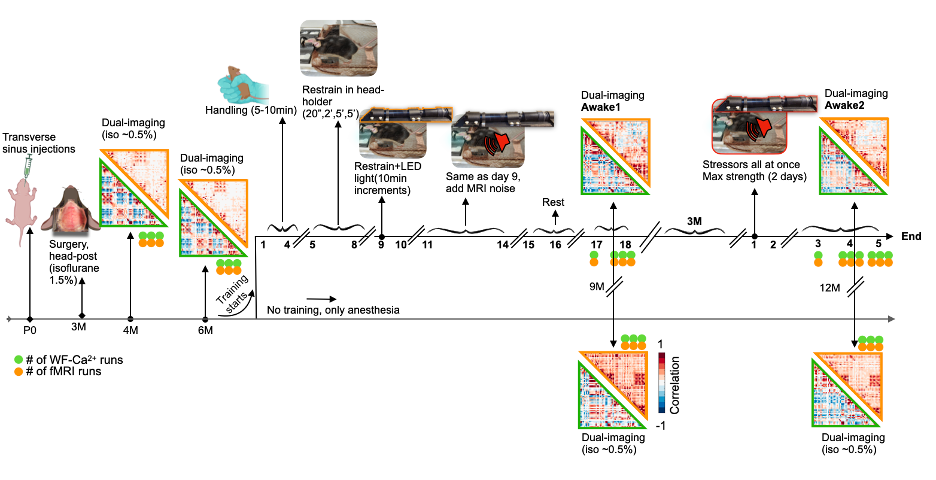
Firstly, N=11 P0 pups undergo transverse sinus injections to encode a GCaMP6s Ca2+ indicator, primarily in excitatory neurons (Fig. 1a,left). At 3 months (M), an optic implant is attached after skull-thinning (Fig. 1a,right). Animals undergo two imaging sessions under isoflurane anesthesia (~0.5%) at 4 and 6M (Fig. 2). Before the 9M timepoint, N=5 mice undergo 14-days of training for awake imaging, where each source of stress (handling, head restrain, LEDs, MRI noise) is gradually introduced. After training, mice are scanned for two consecutive days (Awake1). At 12M, the previously trained mice undergo a refresher training for 2 days where all stressors are provided at max intensity. After the refresher, animals undergo 3 days of imaging (Awake2). Controls are imaged under anesthesia at the same time points. FMRI data are acquired on an 11.7T Bruker scanner, using a gradient-echo, echo-planar-imaging sequence (TR=1.8s, TE=11ms, 0.31mm3 isotropic voxels, 25 slices, 334 volume) with 3 runs/session (indicated with orange dots in Fig. 2). RABIES software is used to preprocess the fMRI data, with frame-to-frame displacement scrubbing threshold at 0.075mm8. WF-Ca2+ data are acquired simultaneously (0.25x0.25μm2) at 10Hz6,7. Data from both modalities are co-registered to a common template for the mouse cortex (Fig. 1d).Results
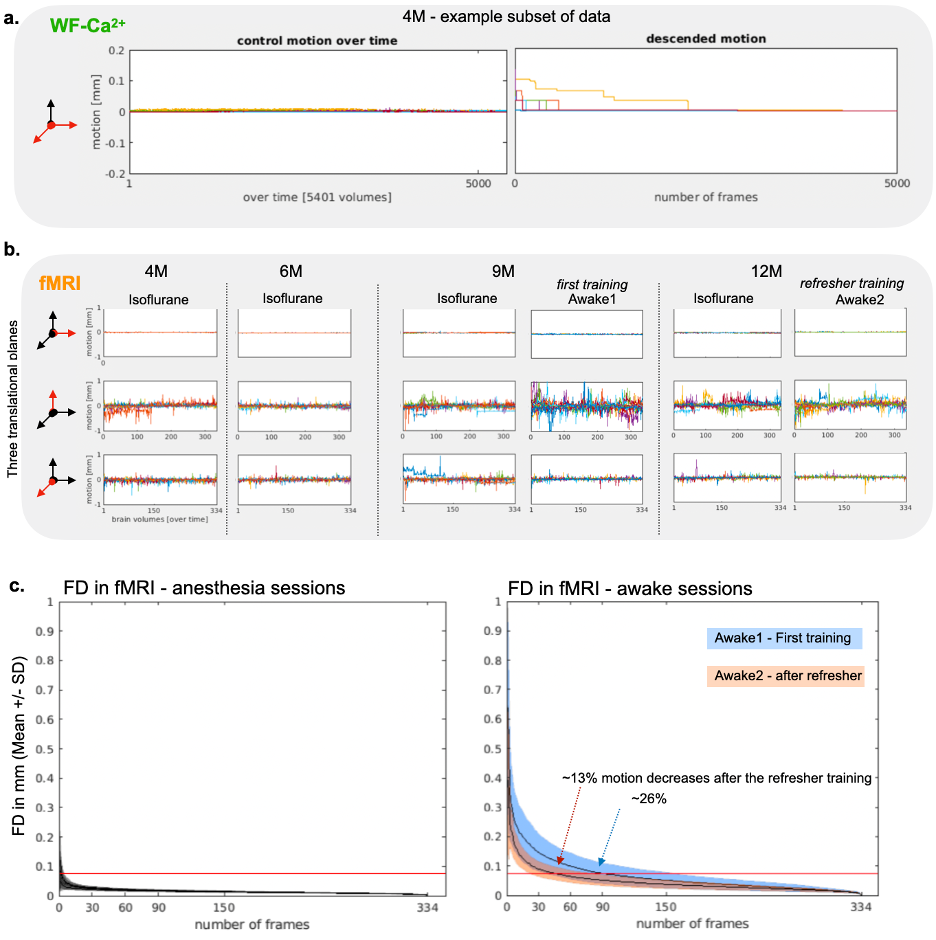
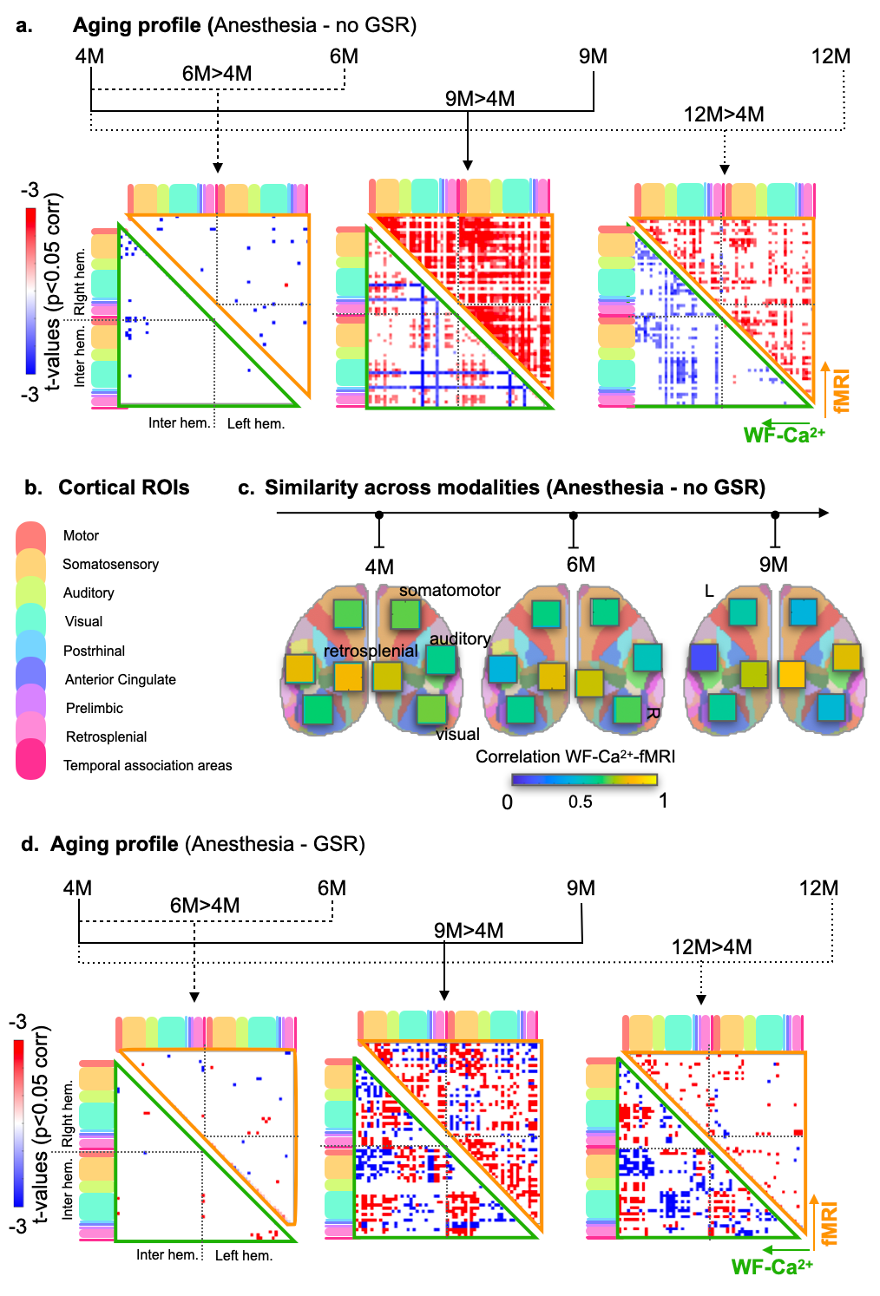
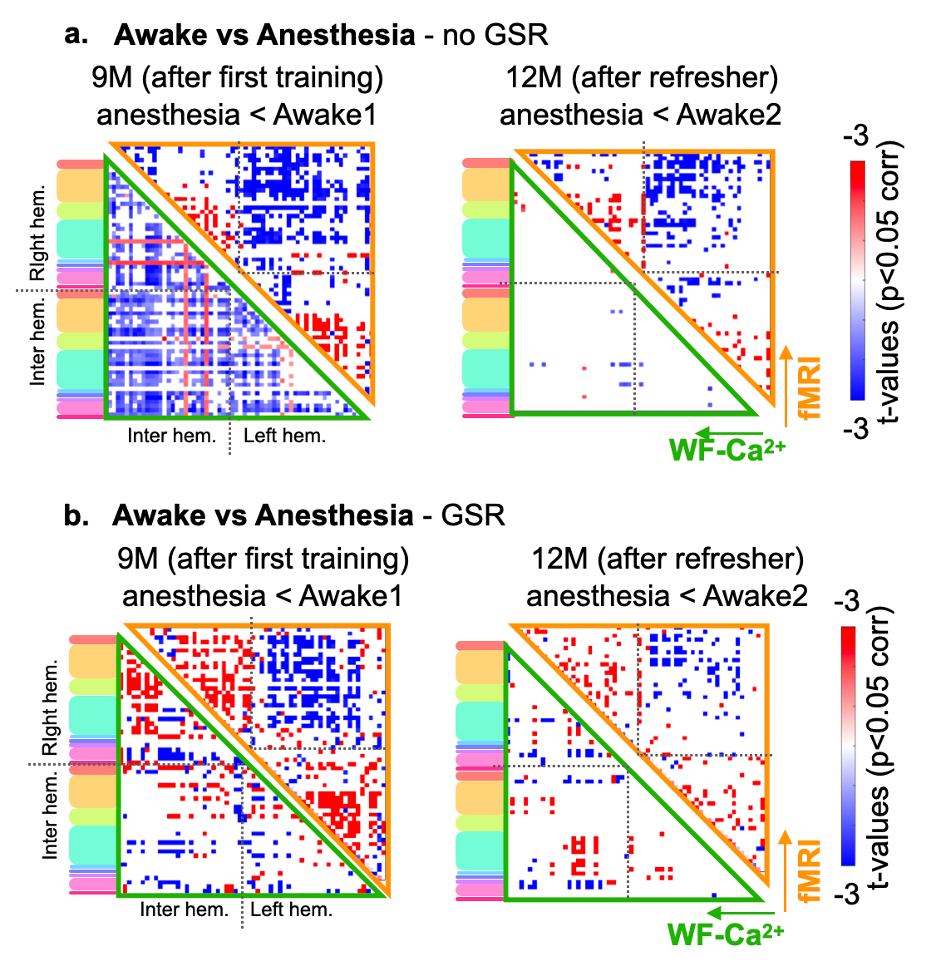
Evidence for implant longevity with example data from fMRI and WF-Ca2+ is shown in Fig. 1b. A 60-ROI atlas is overlayed to extract functional connectomes. Average connectomes from the cortical areas are displayed for all time points/conditions/modalities (Fig. 2, orange/green, fMRI/WF-Ca2+). WF-Ca2+ shows minimal motion (frame-to-frame displacement, FD, over time/high->low, Fig. 3a left/right) in the left-right, anterior-posterior directions. Motion in fMRI data is displayed for all three axes: the superior-inferior direction accounts for the majority of motion across all timepoints (Fig. 3b). The amount of data discarded in the awake session is reduced from 26% (Awake1, in line with previous literature 9-13) to 13% (Awake2), confirming the effectiveness of the refresher in minimizing motion (Fig. 3c). FC analyses in anesthetized animals are reported as t-stat maps for all edges in the 60-ROIs atlas, comparing each timepoint with the earliest one (4M) to assess aging effects. Significant t-values (p<0.05, corrected) are shown as half-matrices both without and with global signal regression (GSR) for both modalities (Fig. 4a,d, green/orange WF-Ca2+/fMRI). An aging effect is reported for both modalities peaking at 9M (Fig. 4a,d). Fig. 4b shows the main regions included in the atlas used. These patterns are confirmed at 12M for the GSR data (Fig. 4d, both modalities), but not entirely for no-GSR data (Fig. 4a) where WF-Ca2+ data show a different trajectory. To explore cross-modality agreement, four areas are selected (example shown for 4, 6, and 9M anesthetized data, no-GSR, Fig. 4c): retrosplenial, visual, auditory, and somatomotor. Some regions show consistent cross-modal agreement, whereas others show disagreement over time (e.g. auditory). Differences in FC in awake vs anesthetized states are shown for both modalities (Fig. 5a,b). Overall, awake fMRI data show a decrease in interhemispheric FC in fMRI, which is recapitulated for no-GSR WF-Ca2+ data at 9M (Fig 5a). The differences in FC between awake and anesthesia (in both modalities) are not as drastic at 12M, although they reveal a similar trajectory.Discussion
We introduce a dual-training program to acclimatize awake mice to fMRI where the second refresher training significantly reduces motion. The combination of fMRI measures with WF-Ca2+ demonstrates that most MR-related motion comes from the superior-inferior direction. Both modalities show an aging effect, although they show some diverging trajectories at later time points, suggesting neurovascular uncoupling. The application of GSR seems to eliminate some important features in the signal. Both modalities show lower inter-hemispheric FC in awake mice vs anesthetized, suggesting that inter-hemispheric connectivity may be enhanced with anesthesia. Results confirm the importance of multi-modal approaches to understanding the complex interplay of neuroimaging signals measuring brain function.Acknowledgements
The staff at Neurotechnology Core (Drs. Joel Greenwood, Paul Shambles and Omer Mano), Dr. Peter Brown from Yale School of Medicine, department of Radiology and biomedical imaging for their technological expertise. Yale Alzheimer’s Disease Research Center Scholar’s Award Funding.References
1.Slupe AM, Kirsch JR. Effects of anesthesia on cerebral blood flow, metabolism, and neuroprotection. J Cereb Blood Flow Metab. 2018 Dec;38(12):2192-2208.
2.Rungta RL, Osmanski BF, Boido D, Tanter M, Charpak S. Light controls cerebral blood flow in naive animals. Nat Commun. 2017 Jan 31;8:14191.
3.Chen X, Tong C, Han Z, Zhang K, Bo B, Feng Y, Liang Z. Sensory evoked fMRI paradigms in awake mice. Neuroimage. 2020 Jan 1;204:116242.
4.Dinh TNA, Jung WB, Shim HJ, Kim SG. Characteristics of fMRI responses to visual stimulation in anesthetized vs. awake mice. Neuroimage. 2021 Feb 1;226:117542.
5.Lake EMR, Ge X, Shen X, Herman P, Hyder F, Cardin JA, Higley MJ, Scheinost D, Papademetris X, Crair MC, Constable RT. Simultaneous cortex-wide fluorescence Ca2+ imaging and whole-brain fMRI. Nat Methods. 2020 Dec;17(12):1262-1271.
6.O'Connor D, Mandino F, Shen X, Horien C, Ge X, Herman P, Hyder F, Crair M, Papademetris X, Lake EM, Constable RT. Functional network properties derived from wide-field calcium imaging differ with wakefulness and across cell type. NeuroImage. 2022 Dec 1;264:119735.
7.Vafaii, H., Mandino, F., Desrosiers-Gregoire, G., O'Connor, D., Shen, X., Ge, X., Herman, P., Hyder, F., Papademetris, X., Chakravarty, M. and Crair, M., 2023. Multimodal measures of spontaneous brain activity reveal both common and divergent patterns of cortical functional organization. Research square, pp.rs-3.
8.Hamodi AS, Martinez Sabino A, Fitzgerald ND, Moschou D, Crair MC. Transverse sinus injections drive robust whole-brain expression of transgenes. Elife. 2020 May 18;9:e53639.
9.Desrosiers-Gregoire G, Devenyi GA, Grandjean J, Chakravarty MM. Rodent Automated Bold Improvement of EPI Sequences (RABIES): A standardized image processing and data quality platform for rodent fMRI. bioRxiv. 2022:2022-08.
10.Harris AP, Lennen RJ, Marshall I, Jansen MA, Pernet CR, Brydges NM, Duguid IC, Holmes MC. Imaging learned fear circuitry in awake mice using fMRI. Eur J Neurosci. 2015 Sep;42(5):2125-34.
11.Madularu D, Mathieu AP, Kumaragamage C, Reynolds LM, Near J, Flores C, Rajah MN. A non-invasive restraining system for awake mouse imaging. J Neurosci Methods. 2017 Aug 1;287:53-57.
12.Han Z, Chen W, Chen X, Zhang K, Tong C, Zhang X, Li CT, Liang Z. Awake and behaving mouse fMRI during Go/No-Go task. Neuroimage. 2019 Mar;188:733-742.
13.Desjardins M, Kılıç K, Thunemann M, Mateo C, Holland D, Ferri CGL, Cremonesi JA, Li B, Cheng Q, Weldy KL, Saisan PA, Kleinfeld D, Komiyama T, Liu TT, Bussell R, Wong EC, Scadeng M, Dunn AK, Boas DA, Sakadžić S, Mandeville JB, Buxton RB, Dale AM, Devor A. Awake Mouse Imaging: From Two-Photon Microscopy to Blood Oxygen Level-Dependent Functional Magnetic Resonance Imaging. Biol Psychiatry Cogn Neurosci Neuroimaging. 2019 Jun;4(6):533-542.
Figures