0332
Investigating hippocampal-cortical and cortical-cortical connectivity changes during pattern separation: a 7T fMRI study1Cleveland Clinic Nevada, Las Vegas, NV, United States, 2Cleveland Clinic Ohio, Cleveland, OH, United States, 3Cleveland Clinic Ohio, Cleveland Clinic, OH, United States, 4University of Colorado, Boulder, Boulder, CO, United States
Synopsis
Keywords: Functional Connectivity, fMRI (task based), pattern separation; hippocampal-cortical connection; cortical-cortical connection
Motivation: Studies have extensively demonstrated roles of hippocampus and its subdivisions during pattern separation, but cortical involvement has not yet been elucidated.
Goal(s): Our goal is to evaluate whole-brain functional connectivity changes during pattern separation.
Approach: We compared cortical-hippocampus and cortical-cortical FNCs during a total of 258 correct and incorrect lure discrimination trials, using high-resolution and high-quality 7T fMRI data.
Results: Cortical-CA3DG FNCs and cortical-CA1 FNCs were significantly involved during pattern separation and completion, respectively. Around 83.35% cortical-cortical connections were with higher FNCs during lure discriminations, indicating their potential involvement during pattern separation.
Impact: Besides hippocampus and its subdivisions, cortical regions and its connections to hippocampus might be extensively involved in pattern separation process.
Background
Pattern separation is a process that discriminates similar object representations in memory[1,2]. Extensive functional MRI (fMRI) studies have focused on the hippocampus and its subdivisions during this process[3–6], with much less attentions being paid to cortical regions. Consequently, roles of interactions between cortical regions with hippocampus and within cortical regions during pattern separation remain unclear. In addition, decreased pattern separation performance has been reported during aging, but how brain functions underlying these changes remain less clear. In this study, we aim to address these questions with whole-brain high-resolution 7T fMRI data during a mnemonic similarity task[2].Methods
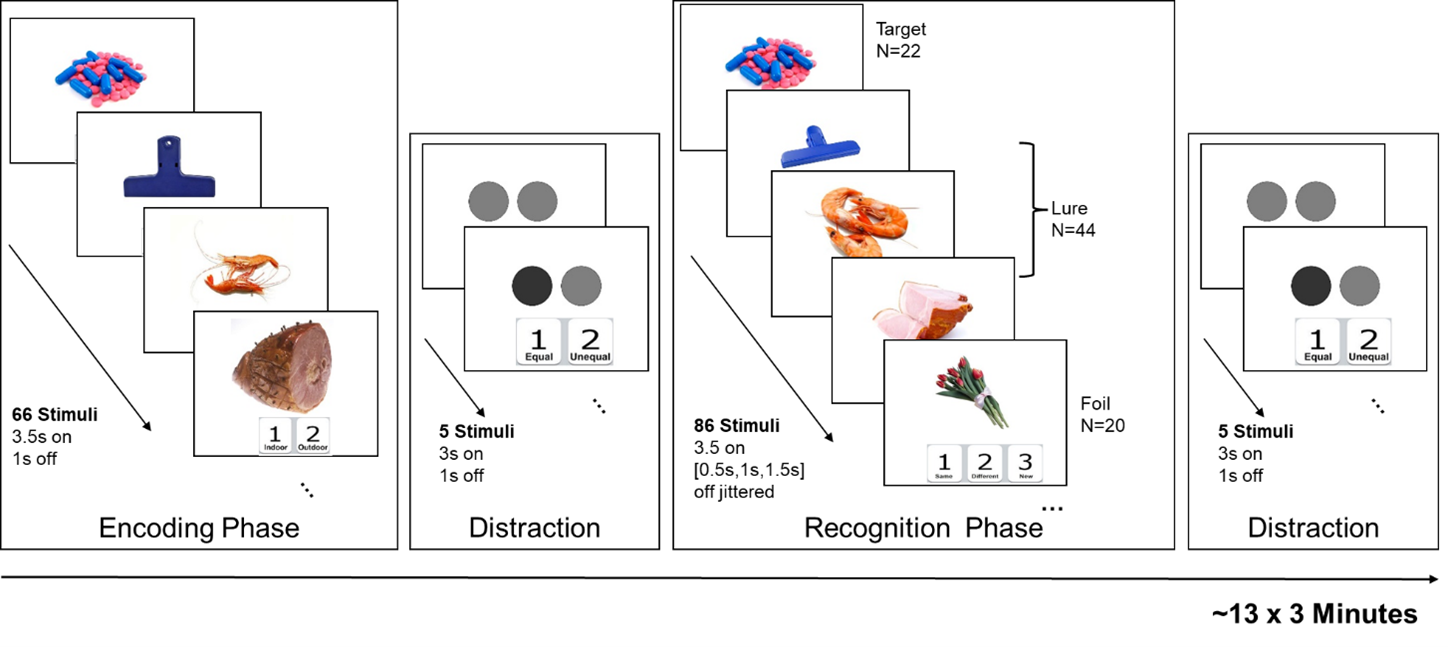
MRI Data collection. MRI data were acquired from 26 non-demented elderly participants on a 7-Tesla Siemens scanner (71.38±4.57 years old, 11 Males/15 Females), including a standard T1-weighted image (voxel-size=0.83x0.83x0.83mm3), a high-resolution T2-weighted image (voxel-size=0.44x0.44x1mm3), and three task-fMRI runs (TR=1.53s, SMS=3, 1.5mm isotropic, 510 time-frames). Fig. 1 depicts the detailed fMRI task design. Briefly, a set of everyday objects were first presented in the encoding phase. The same (targets), similar (lures) and new (foils) objects were later presented in the recognition phase, during which participants were asked to respond whether the objects were the same, different, or new.MRI Data process. The T1 image was input to the FreeSurfer software (v7.2) to generate a subject-specific whole-brain regions-of-interest (ROI) labeling. Then T1 and T2 images were fed into “segmentHA_T2.sh” command to obtain hippocampal subfield ROIs. Following previous pattern separation fMRI studies[6], six hippocampal ROIs were included bilaterally: anterior CA1, anterior subiculum, anterior CA3+DG, posterior CA1, posterior subiculum, and posterior CA3+DG. FMRI data were first slice-timing corrected. All three fMRI runs were realigned and unwrapped together with a voxel-displacement map computed from the GRE field mapping sequences using SPM12. The mean fMRI image was then co-registered to the T1 image using a revised affine transformation with a dilated white-matter (WM) + grey-matter mask (GM) in Advanced Normalization Tools (ANTs) to improve the inter-modality co-registration. Next, the labeling of hippocampal and cortical ROIs in T1 space was transformed to each individual’s fMRI space with the inverse transformation matrix. The mean time-series of all ROIs were then extracted. Motion censoring and nuisance regressions were performed. ROI time series from recognition phase were extracted, demeaned and standardized for each session individually, then concatenated to compute the functional connectivity (FNC) matrices.
FNC computation. We computed FNC matrices for conditions 1) lure stimuli with “different” responses (lure correct rejection (LureCRs)), 2) lure stimuli with “same” responses (lure false alarms (LureFAs)), and 3) target stimuli with “same” responses (Hits), respectively. Stimuli onset for each condition was first convolved with a standard hemodynamic response function, and original time series were weighted by this convolved signal. Pearson’s correlations between weighted time series from pair-wise ROIs were then computed as the FNC values, and a Fisher’s r-to-z transform was performed.
Statistical Analyses. A mixed effect model design was used to evaluate the FNC differences between LureCRs and LureFAs conditions. A greater FNC in LureCRs than in LureFAs indicated that this FNC was involved in the lure discrimination process (i.e., the pattern separation process). In contrast, a greater LureFAs than LureCRs FNC suggested the potential involvement of pattern completion of this FNC. For FNCs with significant LureCRs vs. LureFAs differences (pFDR<0.05), post-hoc association analyses were performed between LureCRs and LureFAs FNCs with behavioral accuracies of LureCRs minus LureFAs, respectively.
Results and Discussion
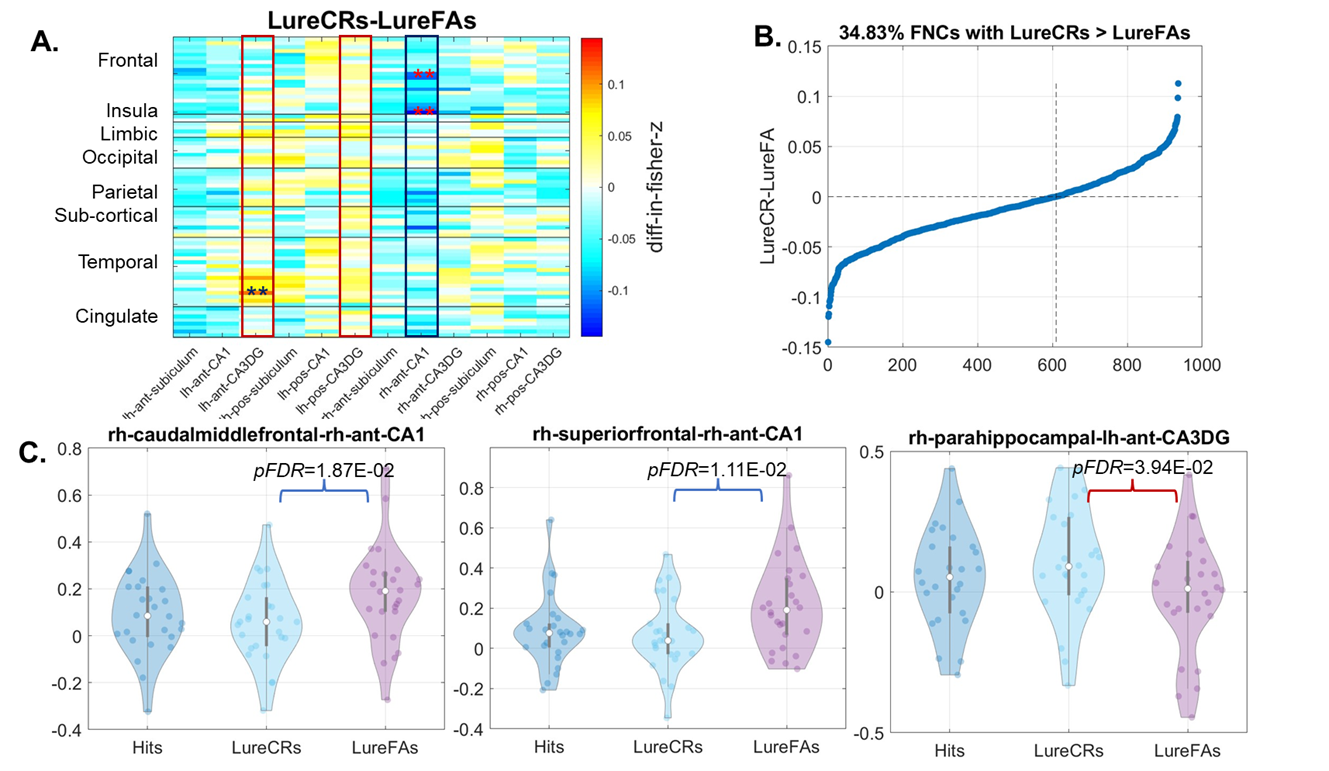
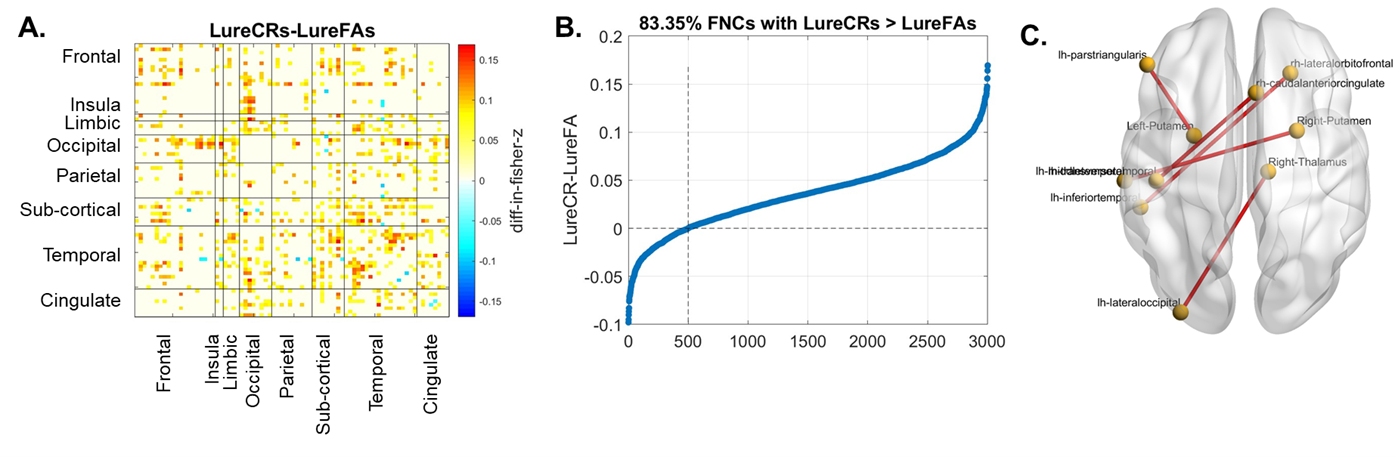
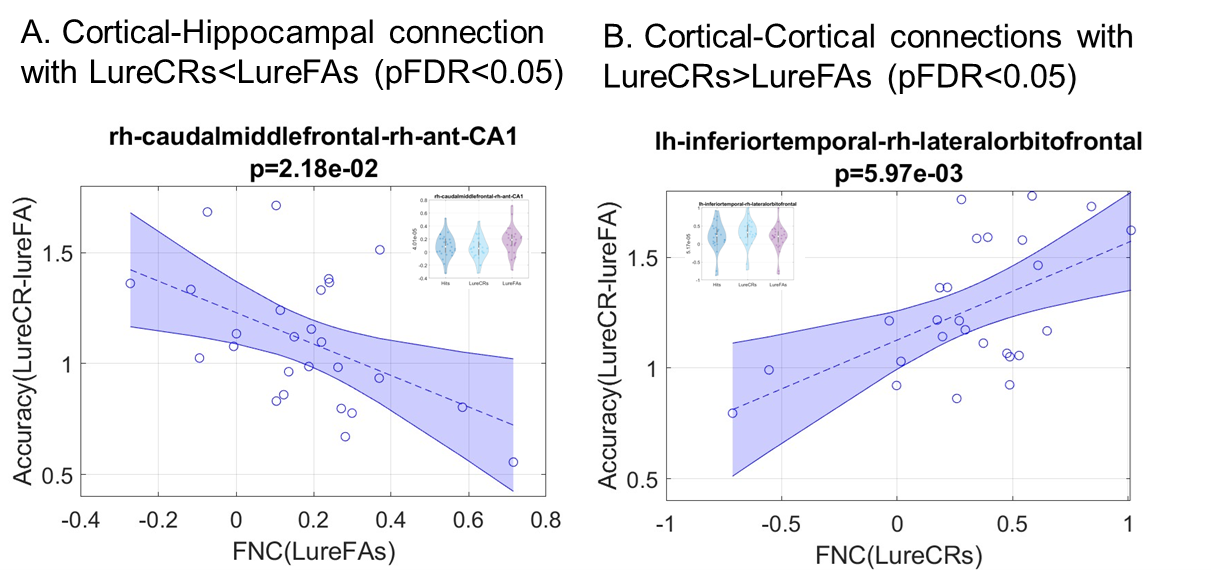
Compared to LureFAs condition, LureCRs condition had greater FNCs for most cortical connections with the left CA3DG area (Fig. 2(A), red boxes), but weaker FNCs for most cortical connections with the CA1 area (Fig. 2(A), blue box). Two frontal-CA1 FNCs (LureFAs>LureCRs) and one temporal-CA3DG FNC (LureFAs<LureCRs) were significantly different (Fig. 2 (C), (pFDR<0.05), which supported the involvement of CA3DG in pattern separation and CA1 in pattern completion. For the frontal-CA1 connection, the negative association between connectivity strength and lure discrimination accuracy further supported this finding (Fig. 4(A)). Interestingly, only 326 out of 936 cortical-hippocampal connections (34.83%, Fig. 2(B)) had greater magnitude during LureCRs than LureFAs conditions. In contrast, 83.35% cortical-cortical connections showed a greater FNC value during LureCRs than LureFAs conditions (Fig. 3(B)), indicating cortical regions might also be extensively involved in the pattern separation process. For significantly different FNCs (Fig. 3(C), pFDR<0.05), a positive association between LureCRs FNCs and lure discrimination accuracies was observed (Fig. 4(B)), further suggested their potential roles in pattern separation.Conclusion
We demonstrated that cortical-CA3DG and cortical-CA1 connections might be separately involved in pattern separation and pattern completion. Our results further indicated that cortical-cortical connections might be extensively involved in the pattern separation process.Acknowledgements
Research reported in this study was supported by NIH RF1AG071566 (NIA), P20GM109025 (NIGMS), P20-AG068053 (NVeADRC). Research reported in thisstudy was additionally supported by private grants from the Peter and Angela Dal Pezzo funds, from Lynn and William Weidner, and from Stacie andChuck Matthewson, and from the Keep Memory Alive Foundation.References
[1] Yassa M, Stark R (2011) Pattern separation in the hippocampus. Trends Neurosci 34, 515–525.
[2] Stark SM, Yassa MA, Lacy JW, Stark CEL (2013) A task to assess behavioral pattern separation (BPS) in humans: Data from healthy aging and mild cognitive impairment. Neuropsychologia 51, 2442–2449.
[3] Reagh ZM, Noche JA, Tustison NJ, Delisle D, Murray EA, Yassa MA (2018) Functional Imbalance of Anterolateral Entorhinal Cortex and Hippocampal Dentate/CA3 Underlies Age-Related Object Pattern Separation Deficits. Neuron 97, 1187-1198.e4.
[4] Yassa MA, Lacy JW, Stark SM, Albert MS, Gallagher M, Stark CEL, Yassa, Michael A, Lacy, Joyce M, Stark, Shauna, Albert, Marilyn, Gallagher Michela, Stark CEL, Yassa MA, Lacy JW, Stark SM, Albert MS, Stark CEL, Gallagher M, Stark CEL, Yassa, Michael A, Lacy, Joyce M, Stark, Shauna, Albert, Marilyn, Gallagher Michela, Stark CEL, Yassa MA, Lacy JW, Stark SM, Albert MS, Stark CEL (2011) Pattern separation deficits associated with increased hippocampal CA3 and dentate gyrus activity in nondemented older adults. Hippocampus 21, 968–979.
[5] Reagh ZM, Yassa MA (2014) Object and spatial mnemonic interference differentially engage lateral and medial entorhinal cortex in humans. Proceedings of the National Academy of Sciences 111, E4264–E4273.
[6] Stevenson RF, Reagh ZM, Chun AP, Murray EA, Yassa MA (2020) Pattern Separation and Source Memory Engage Distinct Hippocampal and Neocortical Regions during Retrieval. J Neurosci 40, 843–851.
Figures