0331
The Role of Inhibitory Thalamic Reticular Nucleus (TRN) in Brain-wide Resting-state Functional MRI (rsfMRI) Connectivity1Laboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Hong Kong SAR, China, 2Department of Electrical and Electronic Engineering, The University of Hong Kong, Hong Kong SAR, China, 3School of Biomedical Sciences, The University of Hong Kong, Hong Kong SAR, China
Synopsis
Keywords: Task/Intervention Based fMRI, fMRI (resting state), fMRI(task based), functional connectivity, neuroscience
Motivation: Despite the enormous potential inherent in rsfMRI, the neural basis underlying rsfMRI connectivity remains unclear.
Goal(s): We aim to dissect the role of the TRN inhibitory neural population in rsfMRI connectivity given its prominent role in maintaining/regulating thalamo-cortical oscillations.
Approach: We examined brain-wide activity and rsfMRI connectivity changes after optogenetically manipulating neural activity in rodent TRN.
Results: We demonstrate that somatosensory-specific TRN inhibitory networks play a role in modulating rsfMRI connectivity of sensorimotor and default mode networks.
Impact: Present studies examining neural basis of rsfMRI have primarily focused on excitatory networks. Here, we investigated the role of a major inhibitory thalamic nucleus to advance our understanding of the contributions of inhibitory inputs in regulating brain-wide rsfMRI networks.
Purpose
It is increasingly clear that resting-state functional MRI (rsfMRI) is a powerful tool for analyzing large-scale functional brain networks. We can now, non-invasively and at the resting state, detect robust and functionally coupled networks in both humans and animals through simple analysis of low frequency temporal oscillations in MRI signals1,2. Thereby, we can interrogate, at a large-scale, whole-brain networks that remain outside of the realm of traditional behavioral/cognitive tasks and neuroimaging approaches. Despite its enormous potential, rsfMRI connectivity remains a correlative/observational measure, because the poorly understood neural basis of rsfMRI poses a barrier for its precise interpretation and effective applications.Our previous rsfMRI studies showed that low frequency (1 Hz) optogenetic stimulation in thalamocortical excitatory neurons in the somatosensory-specific ventral posteromedial (VPM) increased interhemispheric rsfMRI connectivity in all sensory cortices3,4. However, we believe that our findings are likely an oversimplification - when we described the contributions of thalamo-cortical networks in mediating brain-wide rsfMRI connectivity we did so without manipulating and/or monitoring complementary inhibitory activities in the thalamo-cortical circuit.
Here, we propose to dissect the neural basis of rsfMRI connectivity through the thalamic reticular nucleus (TRN), a major source of feedforward and feedback inhibition that executes processing of sensory and associative input to the frontal cortex.
Methods
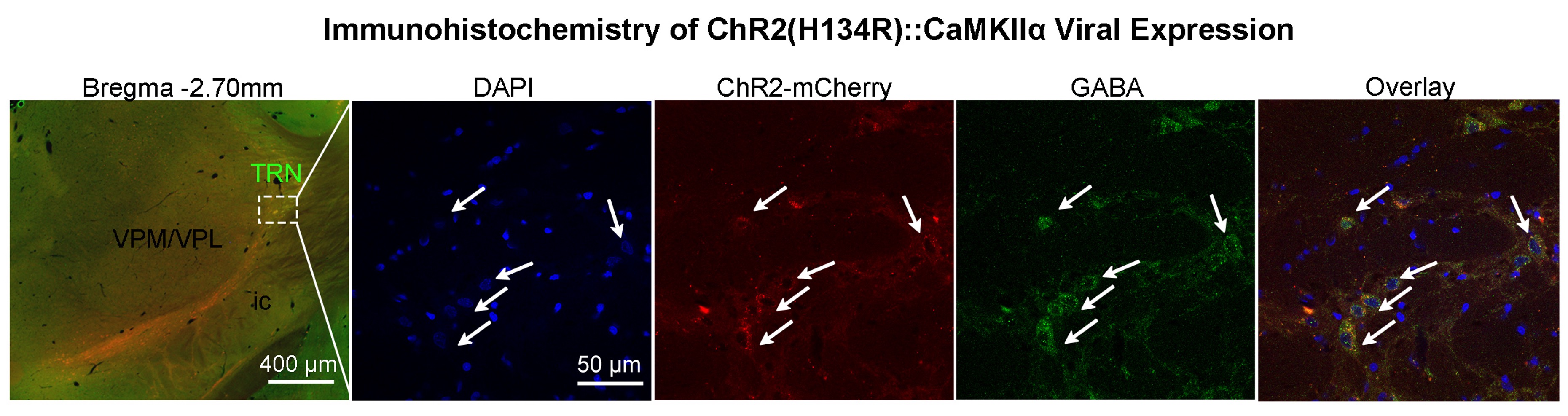
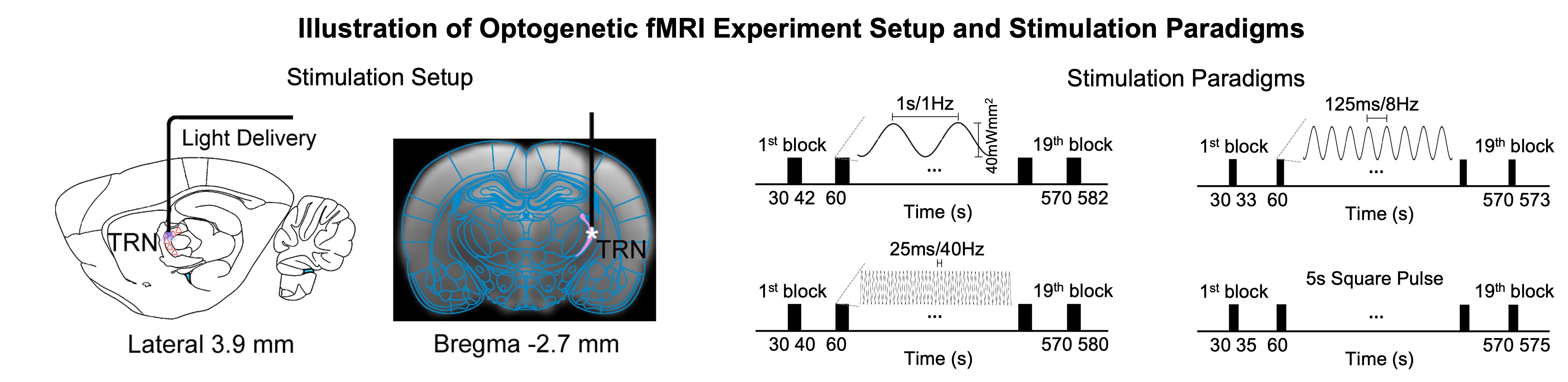
Animal preparation and MRI experimental setup (Figure 1): 3μl rAAV5-hsyn-hChR2(H134R)-mcherry-WPRE-PA was injected to somatosensory-specific subdivision of the TRN in adult SD rats. After 4 weeks, opaque optical fiber cannulas were implanted at the injection sites. All fMRI experiments were performed under 1.0% isoflurane. FMRI data was acquired at 7T using GE-EPI (FOV=32×32mm2, matrix=64×64, α=56°, TE/TR=20/1000ms, and 16 contiguous slices with 1mm thickness).Optogenetic fMRI experiments (Figure 2): Three types of sinusoidal stimuli were implemented to mimic spindle (8Hz, duration=3s, ramp-modulated amplitude), slow (1Hz, duration=12s, constant amplitude) and gamma oscillations (i.e., 40Hz, duration=10s, constant amplitude). An additional long-duration pulse was also presented (duration=5s, constant amplitude) following a paradigm established in a previous work stimulating TRN5. All stimuli were blue (473nm) light with peak amplitudes at 40mW/mm2 and interleaved. Typically, 3-4 baseline rsfMRI scans were acquired before commencement of TRN optogenetic stimulations. GLM analysis was applied to identify significant BOLD responses. Seed-based analysis with task regressors was utilized to quantify rsfMRI connectivity measures during TRN optogenetic stimulation.
Results and Discussion
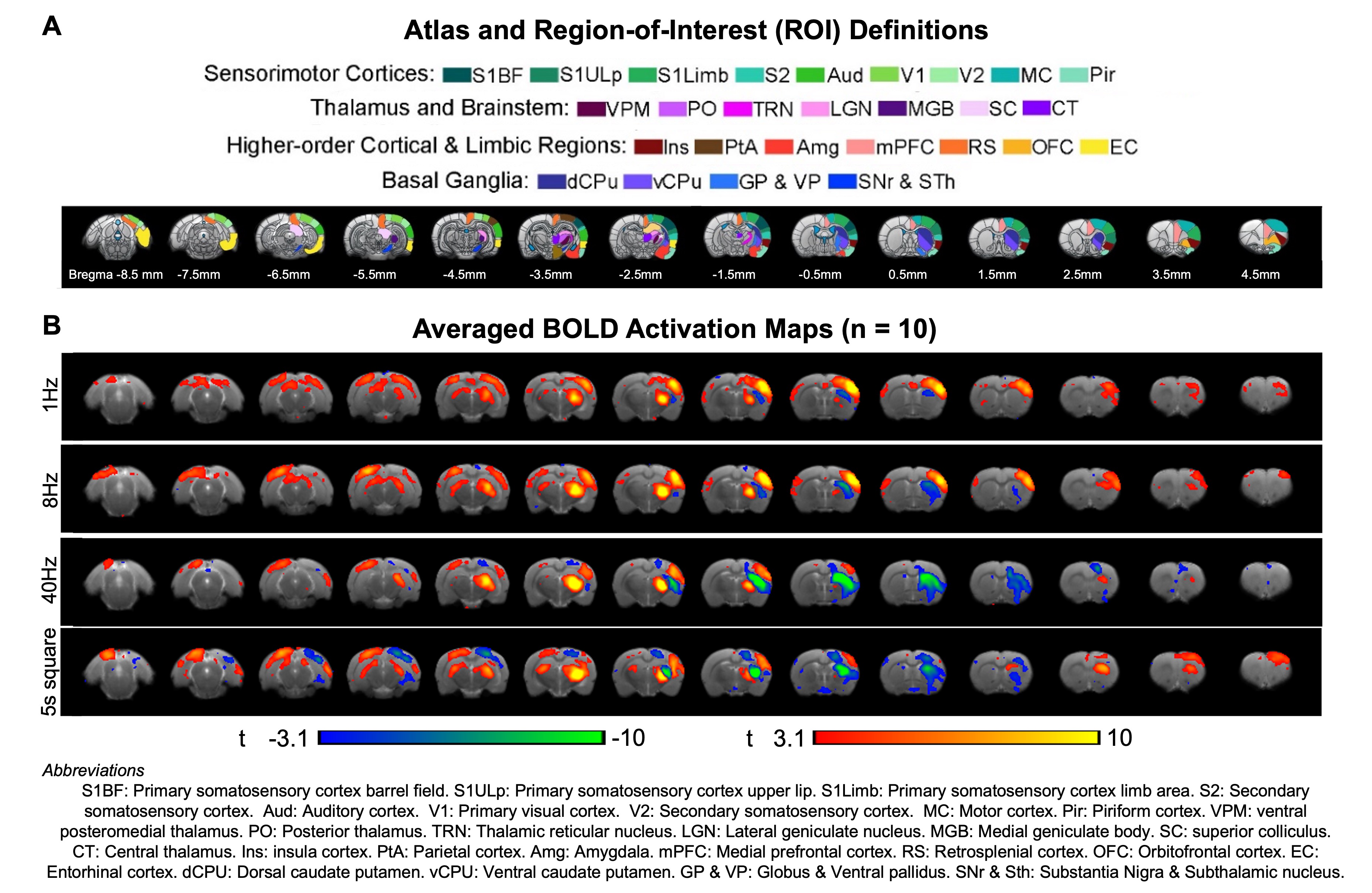
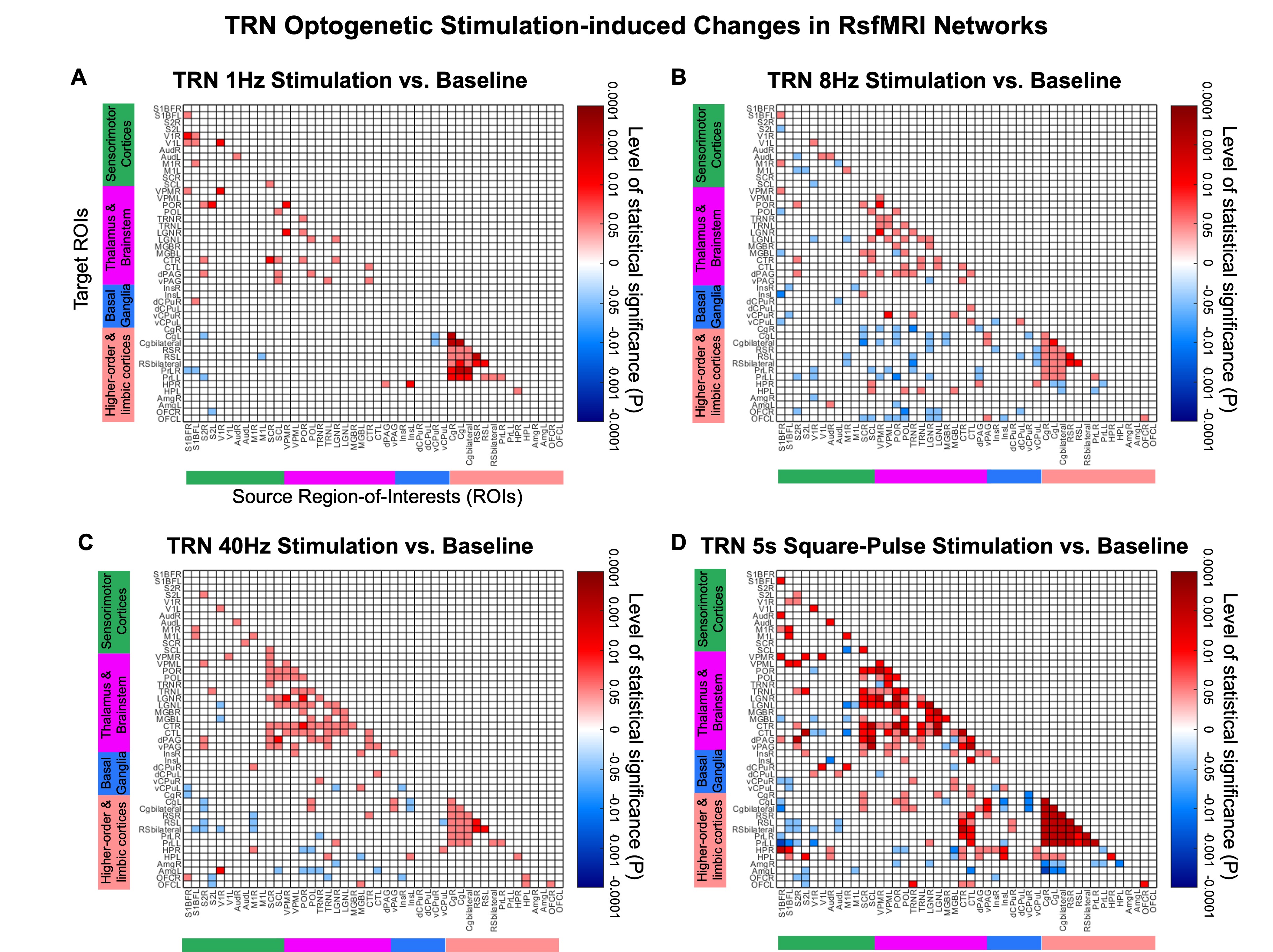
TRN initiated neural oscillations drive distinct brain-wide responses (Figure 3): Optogenetic stimulation of TRN at 1Hz and 8Hz evoked robust positive BOLD responses mainly in ipsilateral sensorimotor cortical and thalamic regions, including somatosensory (primary: barrel field S1BF; limb region S1Limb; upper lip region S1ULp; secondary: S2) and visual (VC) cortices, and somatosensory (ventral posteromedial, VPM) and visual (LGN) thalamus. Meanwhile, at 40Hz, positive BOLD responses were significantly weaker at ipsilateral sensorimotor cortices. Notably, strong negative BOLD responses were evoked at ipsilateral caudate putamen (CPu). In the presence of a long-5s continuous pulse stimulus, strong negative BOLD responses spread to CPu and ipsilateral VC. TRN can drive different oscillations5-7 (slow vs. spindle vs. gamma oscillations) at different levels of synchronization, contributing to the differential fMRI responses ranging from local VPM/S1 responses and brain-wide responses to feedforward inhibition in remote regions, demonstrating the role of TRN in regulating inhibition-excitation balance in thalamo-cortical networks.TRN modulates brain-wide rsfMRI networks (Figure 4): RsfMRI connectivity in bilateral sensorimotor cortices and thalamic nuclei, and within default mode network (DMN; comprising of higher-order and limbic cortices ROIs) were consistently increased in all TRN stimulation paradigms. Interestingly, we observed specific deviations in the modulatory effects on rsfMRI connectivity. For example, connectivity from thalamus & brainstem to DMN was decreased during 8Hz but increased during 5s square-pulse stimulations. Given the dense reciprocal projections between the excitatory and inhibitory neural populations in VPM and TRN8,9 and that dysfunction in the thalamic reticular nucleus (TRN) significantly disrupts thalamo-cortical oscillations6,10, we hypothesize that an overall changes in rsfMRI connectivity is likely mediated by reciprocal projections between both excitatory and inhibitory neural populations in the VPM and the TRN, respectively.
Conclusion
In this study, we investigated the neural basis of rsfMRI connectivity through the thalamic reticular nucleus (TRN). We found that TRN and its inhibitory networks play a central role in modulating brain-wide functional networks as measured with rsfMRI. Although our present findings revealed varied rsfMRI connectivity changes during different stimulation paradigms, subsequent electrophysiological recordings are needed at the identified key ROIs studies to elucidate the underlying neural events that are associated with the altered rsfMRI networks.Acknowledgements
This work was supported in part by Hong Kong Research Grant Council (HKU17112120, HKU17127121, HKU17127022 and HKU17127523 to E.X.W.; and HKU17104020, HKU17127021, HKU17127723 to A.T.L.L.), Lam Woo Foundation, and Guangdong Key Technologies for AD Diagnostic and Treatment of Brain (2018B030336001) to E.X.W.References
- Miller, K.L., et al. Multimodal population brain imaging in the UK Biobank prospective epidemiological study. Nat Neurosci 19, 1523-1536 (2016).
- Glasser, M.F., Smith, S.M., Marcus, D.S., Andersson, J.L.R., Auerbach, E.J., Behrens, T.E.J., Coalson, T.S., Harms, M.P., Jenkinson, M., Moeller, S., Robinson, E.C., Sotiropoulos, S.N., Xu, J., Yacoub, E., Ugurbil, K. & Van Essen, D.C. The Human Connectome Project’s neuroimaging approach. Nature Neuroscience 19, 1175 (2016).
- Leong, A.T.L., Chan, R.W., Gao, P.P., Chan, Y.S., Tsia, K.K., Yung, W.H. & Wu, E.X. Long-range projections coordinate distributed brain-wide neural activity with a specific spatiotemporal profile. Proc Natl Acad Sci U S A 113, E8306-E8315 (2016).
- Wang, X., Leong, A.T.L., Chan, R.W., Liu, Y. & Wu, E.X. Thalamic low frequency activity facilitates resting-state cortical interhemispheric MRI functional connectivity. Neuroimage 201, 115985 (2019).
- Lewis, L.D., Voigts, J., Flores, F.J., Schmitt, L.I., Wilson, M.A., Halassa, M.M. & Brown, E.N. Thalamic reticular nucleus induces fast and local modulation of arousal state. eLife 4, e08760 (2015).
- Fogerson, P.M. & Huguenard, J.R. Tapping the Brakes: Cellular and Synaptic Mechanisms that Regulate Thalamic Oscillations. Neuron 92, 687-704 (2016).
- Fernandez, L.M., Vantomme, G., Osorio-Forero, A., Cardis, R., Beard, E. & Luthi, A. Thalamic reticular control of local sleep in mouse sensory cortex. eLife 7, e39111 (2018).
- Pinault, D. The thalamic reticular nucleus: structure, function and concept. Brain Res Brain Res Rev 46, 1-31 (2004).
- Li, Y., et al. Distinct subnetworks of the thalamic reticular nucleus. Nature (2020).
- Krol, A., Wimmer, R.D., Halassa, M.M. & Feng, G. Thalamic Reticular Dysfunction as a Circuit Endophenotype in Neurodevelopmental Disorders. Neuron 98, 282-295 (2018).
Figures