0329
Structural and functional connectivity patterns of brainstem nuclei in living humans by 7 Tesla MRI1Brainstem Imaging Laboratory, Department of Radiology, Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital and Harvard Medical School, Charlestown, MA, United States, 2Multiscale Imaging and Integrative Biophysics Unit, LBN, National Institute on Aging, NIH, Baltimore, MD, United States, 3Escuela Nacional de Estudios Superiores Unidad Juriquilla, Universidad Nacional Autónoma de México, Querétaro, Juriquilla, Mexico, 4Parkinson's Disease and Movement Disorders Unit, Center for Rare Neurological Diseases (ERN-RND), Department of Neurosciences, University of Padova, Padova, Italy, 5Division of Sleep Medicine, Harvard University, Boston, MA, United States
Synopsis
Keywords: Functional Connectivity, High-Field MRI, Neuro, Structural connectivity, Functional connectivity
Motivation: A definitive baseline connectome of brainstem nuclei is missing.
Goal(s): To improve brainstem hodology in living humans by using the similarity between functional and structural connectomes of brainstem nuclei as ground truth.
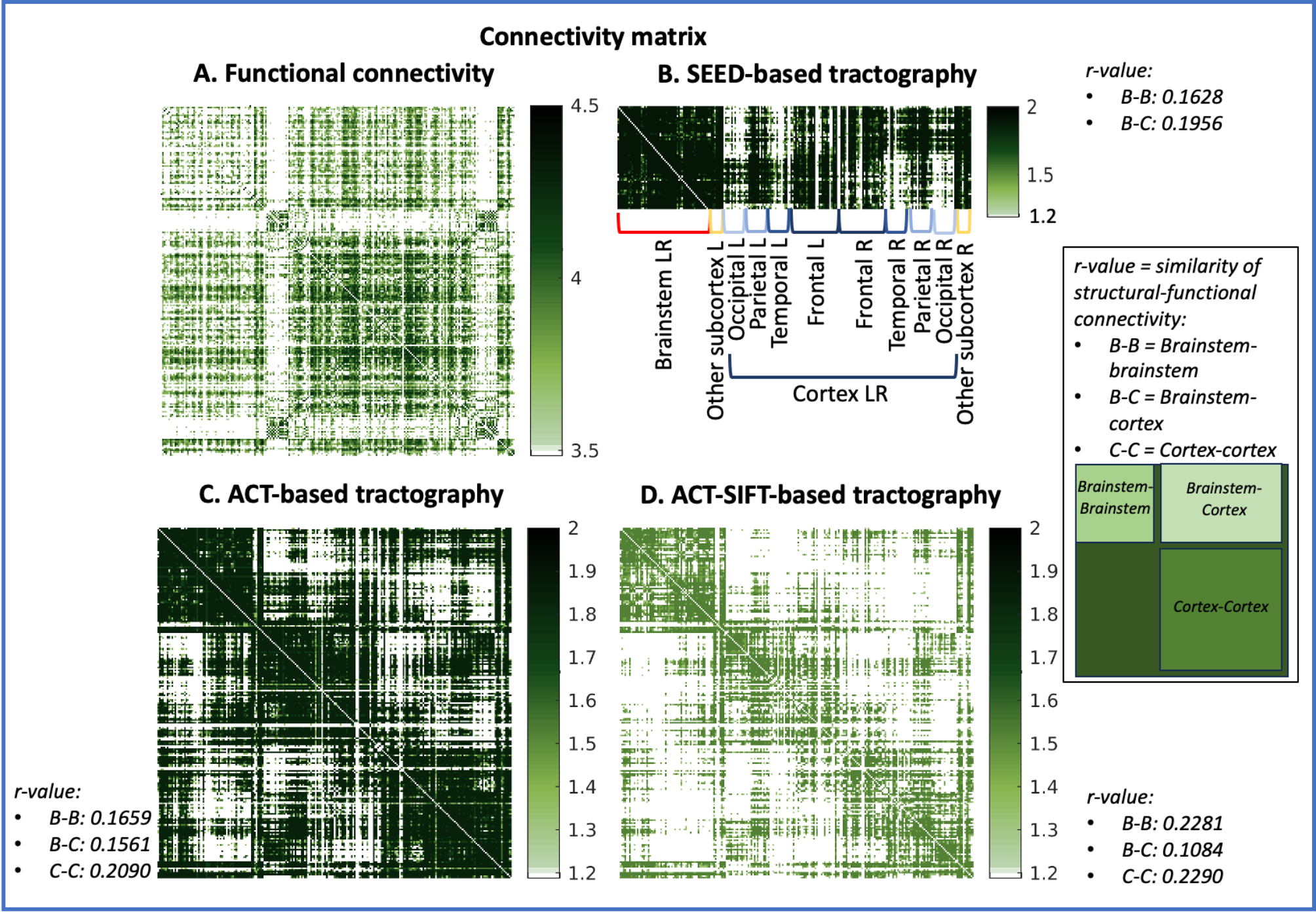
Approach: In healthy subjects, we mapped 58 Brainstem Navigator atlas labels to high spatial resolution functional and diffusion-weighted 7 Tesla MRI, and computed their functional and structural connectivity, the latter computed using three probabilistic tractography methods proposed in the literature (seed-, ACT-, ACT-SIFT-based), with 148 cortical and 21 subcortical areas.
Results: ACT-SIFT outperformed the other methods within the brainstem and the cortex by reducing large fiber bias.
Impact: Comparison of structural and functional connectomes achieved with different methodology can improve the understanding and mapping of brainstem nuclei connections in living humans and establish a baseline connectome useful to evaluate a broad set of diseases including movement/sleep disorders.
Introduction:
Identification of brainstem nuclei pathways (hodology) in living humans is crucial to understand brainstem-related disease, such as movement/sleep/arousal/autonomic disorders. Yet, in-vivo brainstem hodology is currently understudied1-4 due to the small size and deep location of these nuclei; moreover, functional1,2 and structural3,4 brainstem nuclei connectomes derived from in-vivo functional-MRI (fMRI) and diffusion-based-tractography respectively, show similarities but also some discrepancies. For example, the structural connecome is denser within the brainstem, while the functional connectome is denser within the cortex. Thus, there is a crucial unmet need to establish a definitive baseline connectome of brainstem nuclei and understand if these differences are due to sensitivity issues/bias of either technique or are true.Purpose:
To improve brainstem hodology in living humans by using as ground truth the similarity between the functional and structural connectome of brainstem nuclei, the latter computed using three probabilistic tractography methods proposed in the literature.5-7Methods:
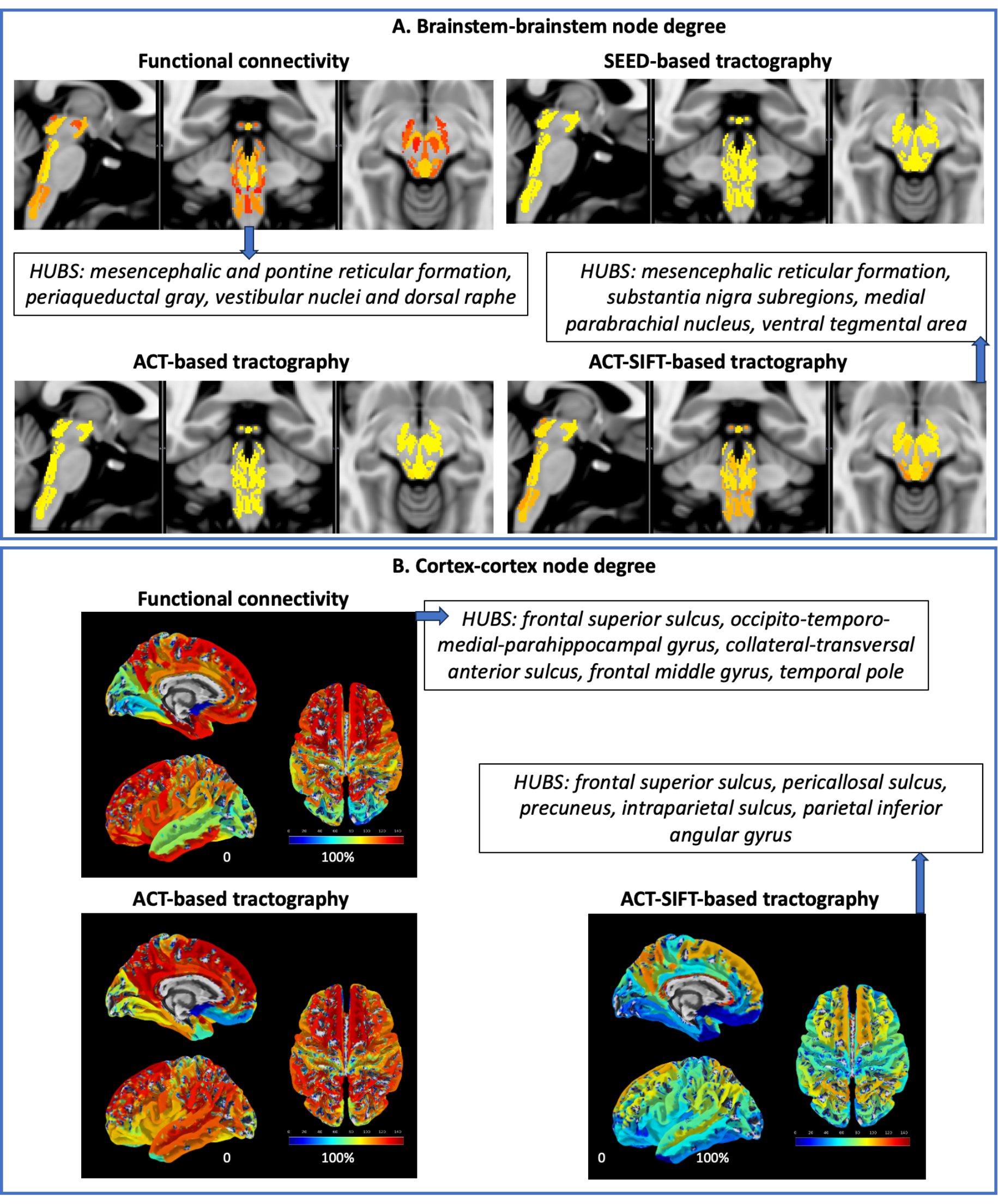
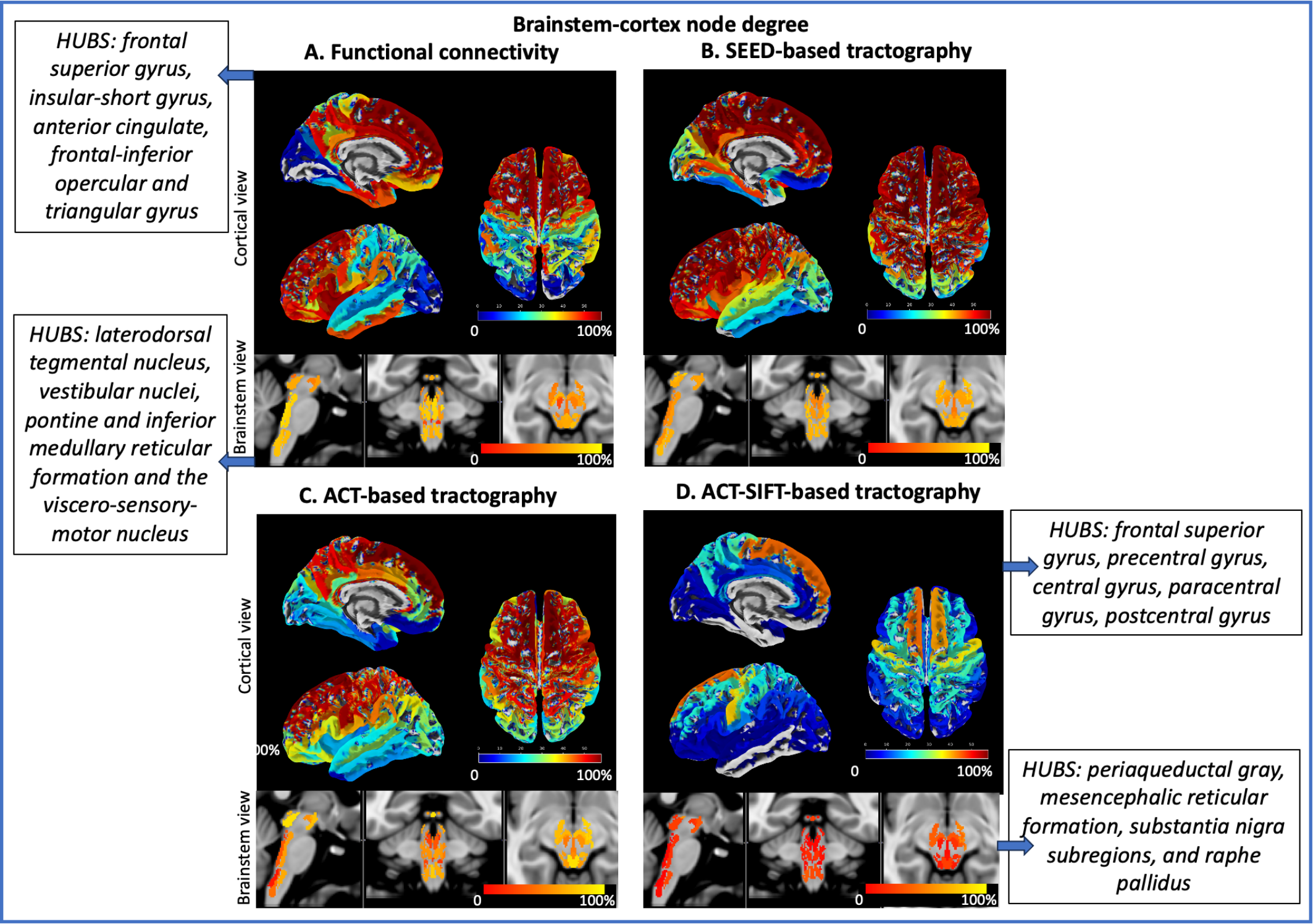
Data acquisition: Twenty healthy volunteers (10m/10f; age 29.5±1.1years) underwent 7 Tesla MRI under IRB approval; results from eight subjects are presented here. fMRI: 3 resting-state (eyes-closed) runs, gradient-echo EPI, isotropic voxel-size/matrix-size/GRAPPA-factor/nominal echospacing/bandwidth/N. slices/slice orientation/slice-acquisition order/echo-time (TE)/repetition-time (TR)/flip-angle(FA)/simultaneous-multi-slice factor/N. repetitions/phase-encoding direction/acquisition-time per-run=1.1mm/180x240/3/0.82ms/“1488Hz/Px”/123/sagittal/interleaved/32ms/2.5s/75°/3/210/anterior-posterior/10’07”. A field-map and a multi-echo MPRAGE (MEMPRAGE) were also acquired. Diffusion-weighted-imaging (DWI): 2D spin-echo EPI, n. slices/echo-time/repetition-time/phase-encoding direction/bandwidth/partial-Fourier/n. diffusion-directions/b-value: 82/66.8ms/7.4s/“anterior/posterior”/“1456Hz/pixel”/“6/8”/60/2500s/mm2, seven interspersed “b0” images, acquisition-time: 8′53′′. Seven “b0” images with posterior/anterior phase-encoding direction were also acquired. Data analysis: a) Nodes used in connectivity analysis: we used 58 brainstem nuclei labels from the Brainstem Navigator8-13; 21 subcortical labels8,14-15, and 148 Freesurfer cortical parcellations15. b) fMRI1-2: images were RETROICOR and slice-timing corrected, reoriented to “RPI”, cropped, distortion corrected using the field-map, motion corrected, coregistered to the MEMPRAGE. Then, we regressed out nuisance time-series due to motion, cardiac-rate and respiratory-volume-per-unit-time fluctuations, and signals in the cerebrospinal fluid neighboring the brainstem. We scaled the signal to percent signal change, removed the temporal mean, and performed band-pass filtering (cut-off 0.01-0.1 Hz). We concatenated the runs and applied the MEMPRAGE-to-stereotactic-space transformations. Finally, we computed the Pearson’s correlation coefficient at the subject level, between average time-courses extracted from nodes. At the group level, we performed a one-sample t-test on the Fisher-transformed correlation coefficients and defined as ‘links’ significant connections (p<0.0005 FDR-corrected). c) DWI: DWIs were denoised, motion- and distortion-corrected. Nodes were aligned to native space from stereotactic space using nonlinear registrations, and three probabilistic tractography methods (all based on the MRtrix3 iFOD2 constrained spherical deconvolution algorithm) were used: 1) SEED-based tractography3-4: this method propagates 100,000 streamlines from within each gray matter node (i.e. seed); note that it was computed only for brainstem nodes; 2) Anatomically constrained tractography (ACT6), which uses anatomical priors and propagates 100 million streamlines from the whole-brain gray-white matter border, enforces streamline termination also at the gray-white matter border, and removes indirect connectivity through subcortical regions; 3) ACT followed by Spherical-deconvolution Informed Filtering of Tractograms (SIFT7), which, beyond ACT features, reduces the tractography bias towards larger fiber bundles (substantially present in the brainstem) by filtering the streamlines to achieve a density matched to the fiber-orientation-distribution lobe integral. For each of these methods, we then counted the fraction of streamlines connecting pairs of nodes, averaged it across subjects and performed a Wilcoxon test (p<0.05 FDR-corrected). Node degree was computed as the number of significant connections of a node within specific connectivity submatrices.Results:
Functional and structural connectivity matrices of brainstem nuclei with the whole brain are shown in Figure 1. Note the stronger functional and structural connectivity of brainstem nuclei with frontal cortex and weaker connectivity with visual cortex. Node degree computed on brainstem-brainstem and cortex-cortex submatrices along with hubs are shown in Figure 2, and on brainstem-cortex submatrix is shown in Figure 3.Discussion and Conclusions:
Interestingly, due to more biologically realistic constraints on streamline seeding/termination (ACT) and reduction of large fiber bias (SIFT), ACT-SIFT improved the similarity of functional and structural connectivity for brainstem-brainstem and cortex-cortex submatrices. Its decreased performance in brainstem-cortex evaluation is most likely related to ACT-SIFT removal of indirect connectivity through subcortex, which is present in functional connectivity, as well as in SEED-based tractography. We are currently evaluating whether partial correlation and other methods17 that account for indirect functional connectivity can further improve the functional-structural connectivity similarity. Comparison of structural and functional connectomes achieved with different methodology can improve the understanding and mapping of brainstem nuclei connections in living humans and establish a baseline connectome useful to evaluate a broad set of brainstem-related diseases.Acknowledgements
NIH NIA R01 AG063982; Dr. Thorsten Feiweier for providing the diffusion sequence used in this study.References
1. Cauzzo S, Singh K, Stauder M, et al. Functional connectome of brainstem nuclei involved in autonomic, limbic, pain and sensory processing in living humans from 7 Tesla resting state fMRI. Neuroimage. 2022;250:118925.
2. Singh K, Cauzzo S, García-Gomar MG, et al. Functional connectome of arousal and motor brainstem nuclei in living humans by 7 Tesla resting-state fMRI. NeuroImage. 2022;249:118865.
3. Singh K, García‐Gomar MG, Cauzzo S, et al. Structural connectivity of autonomic, pain, limbic, and sensory brainstem nuclei in living humans based on 7 Tesla and 3 Tesla MRI. Human Brain Mapping. 2022;43(10):3086-3112.
4. García‐Gomar MG, Singh K, Cauzzo S, et al. In vivo structural connectome of arousal and motor brainstem nuclei by 7 Tesla and 3 Tesla MRI. Human Brain Mapping. 2022;43(14):4397-4421.
5. Tournier JD, Calamante F, Connelly A. MRtrix: Diffusion tractography in crossing fiber regions. Int J Imag Syst Tech. 2012; 22(1):53-66.
6. Smith RE, Tournier JD, Calamante F, et al. Anatomically-constrained tractography: improved diffusion MRI streamlines tractography through effective use of anatomical information. Neuroimage. 2012;62(3):1924-1938.
7. Smith RE, Tournier JD, Calamante F, et al. SIFT: Spherical-deconvolution informed filtering of tractograms. Neuroimage. 2013;67:298-312.
8. Bianciardi M, Toschi N, Edlow BL, et al. Toward an in vivo neuroimaging template of human brainstem nuclei of the ascending arousal, autonomic, and motor systems. Brain Connect. 2015;5(10):597-607.
9. Bianciardi M, Strong C, Toschi N, et al. A probabilistic template of human mesopontine tegmental nuclei from in vivo 7 T MRI. Neuroimage. 2018;170:222-230.
10. García-Gomar MG, Strong C, Toschi N, et al. In vivo probabilistic structural atlas of the inferior and superior colliculi, medial and lateral geniculate nuclei and superior olivary complex in humans based on 7 tesla MRI. Front Neurosci. 2019;13:764.
11. Singh K, Indovina I, Augustinack JC, et al. Probabilistic template of the lateral parabrachial nucleus, medial parabrachial nucleus, vestibular nuclei complex, and medullary viscero-sensory-motor nuclei complex in living humans from 7 Tesla MRI. Front Neurosci. 2020;13:1425.
12. Singh K, García-Gomar MG, Bianciardi M. Probabilistic atlas of the mesencephalic reticular formation, isthmic reticular formation, microcellular tegmental nucleus, ventral tegmental area nucleus complex, and caudal–rostral linear raphe nucleus complex in living humans from 7 Tesla magnetic resonance imaging. Brain Connect. 2021;11(8):613-623.
13. García-Gomar MG, Videnovic A, Singh K, et al. Disruption of Brainstem Structural Connectivity in REM Sleep Behavior Disorder Using 7 Tesla Magnetic Resonance Imaging. Mov Disord. 2022; 37(4):847-853.
14. Pauli WM, Nili AN, Tyszka JM. A high-resolution probabilistic in vivo atlas of human subcortical brain nuclei. Scientific Data. 2018;5:180063.
15. Destrieux C, Fischl B, Dale A, et al. Automatic parcellation of human cortical gyri and sulci using standard anatomical nomenclature. NeuroImage. 2010;53(1):1–15.
16. Buckner RL, Sepulcre J, Talukdar T, et al. Cortical hubs revealed by intrinsic functional connectivity: mapping, assessment of stability, and relation to Alzheimer's disease. J Neurosci. 2009;29(6):1860-73.
17. Sundaram P, Luessi M, Bianciardi M, et al. Individual resting-state brain networks enabled by massive multivariate conditional mutual information. IEEE transactions on medical imaging. 2019;39(6):1957-1966.
Figures