0212
Astrocytes contribute to signals of Hyperpolarized 13C pyruvate in the brain1Institute for Quantum Life Science, National Institutes for Quantum Science and Technology, Chiba, Japan, 2RIKEN Center for Brain Science, Wako, Japan, 3Department of Advanced Nuclear Medicine Sciences, Institute for Quantum Medical Science, National Institutes for Quantum Science and Technology, Chiba, Japan, 4Bruker Japan K.K., Yokohama, Japan, 5National Institutes for Quantum Science and Technology, Chiba, Japan
Synopsis
Keywords: Hyperpolarized MR (Non-Gas), Hyperpolarized MR (Non-Gas)
Motivation: Hyperpolarized MR (HP-MR) 13C pyruvate is a valuable probe for evaluating glycolytic flux, but it remains unknown which cells contribute to the HP-MR signals in the brain.
Goal(s): To verify whether astrocyte metabolism is involved in the signal of HP-MR 13C pyruvate in the brain.
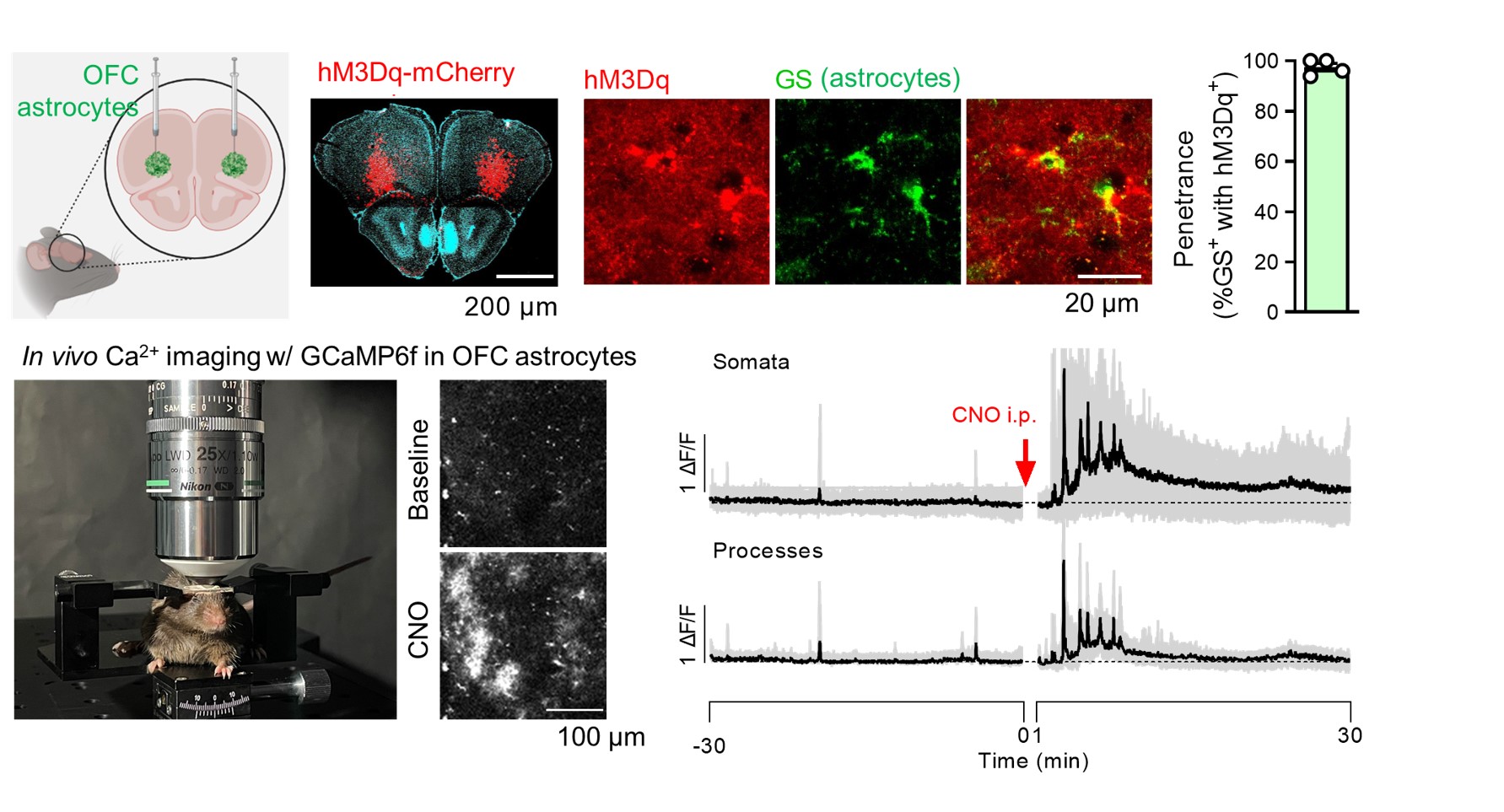
Approach: We perturbed Gq-GPCR/calcium (Ca2+) signaling in astrocytes using the hM3Dq DREADD system and examined the fluctuations of the HP-MR pyruvate signals in awake mice.
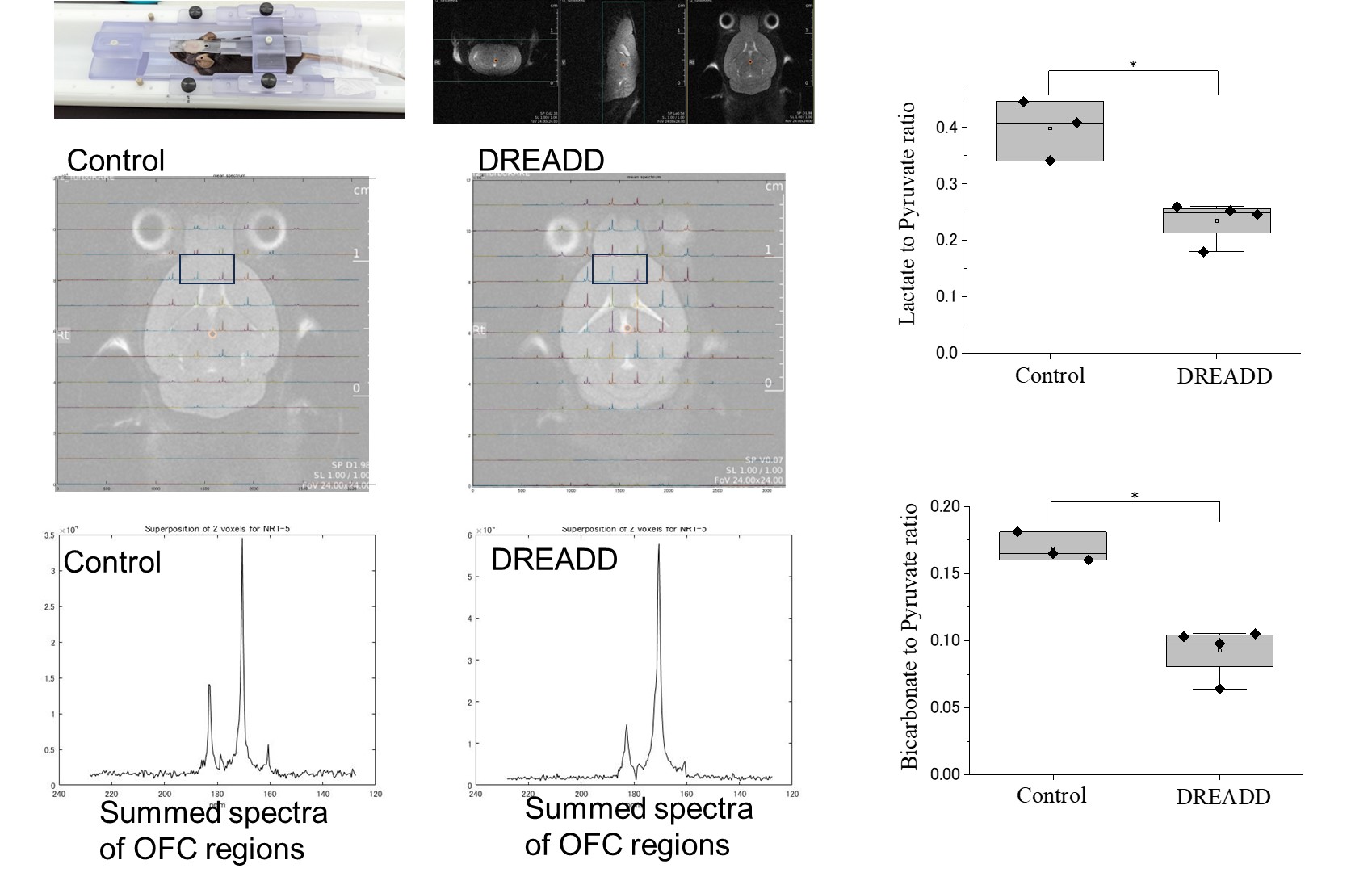
Results: In HP-MR experiments conducted 30 minutes after the elevation of astrocyte Ca2+, the lactate/pyruvate ratio decreased compared to the control, and the bicarbonate/pyruvate ratio also decreased.
Impact: Investigating the involvement of astrocyte metabolism in brain hyperpolarized MR(HP-MR) 13C pyruvate signals, astrocyte GPCR/Ca2+ signaling was perturbed using DREADD, revealing a decrease in lactate/pyruvate and bicarbonate/pyruvate ratios in HP-MR experiments, suggesting metabolic alterations in response to astrocyte GPCR/Ca2+ modulation.
INTRODUCTION
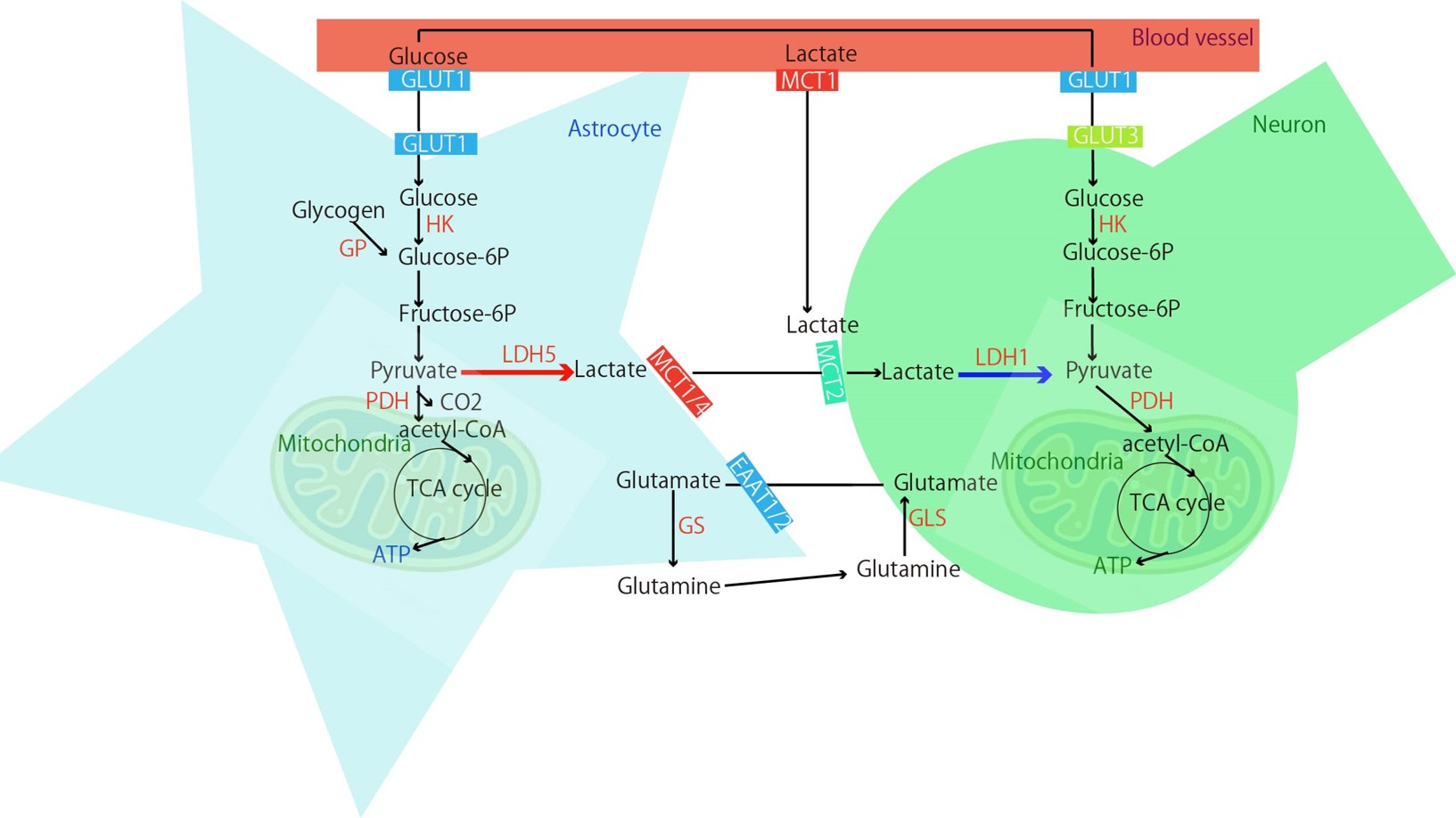
Hyperpolarized 13C-MRI (HP-MR) is a technology that enables the visualization of the conversion from 1-13C pyruvate to 1-13C lactate by dramatically increasing the measurement sensitivity of NMR. The glycolytic pathway is suggested to be abnormal not only in cancer but also in brain disorders, including Alzheimer's disease1. Both neurons and astrocytes contribute to the regulation of brain energy metabolism. A hypothesis has also been proposed that astrocytes take up glucose from blood vessels, convert it to pyruvate, then to lactate, and that lactate is used by astrocytes2 (Astrocyte-Neuron Lactate Shuttle, ANLS). Given the crosstalk between neurons and astrocytes, the conversion from pyruvate to lactate might be mainly conducted by astrocytes. However, such verification has not been performed with HP-MR for converting from 1-13C pyruvate to lactate in the living brain. We manipulated the calcium ion (Ca2+) of astrocytes using the DREADD system (Designer Receptors Exclusively Activated by Designer Drugs)3 and varied the metabolic state, and verified the contribution of astrocyte activity to HP-MR by measuring brain metabolism with HP-MR. In parallel, verification was also performed with 18F-FDG-PET, for which the contribution of astrocytes has been reported4,5.METHOD
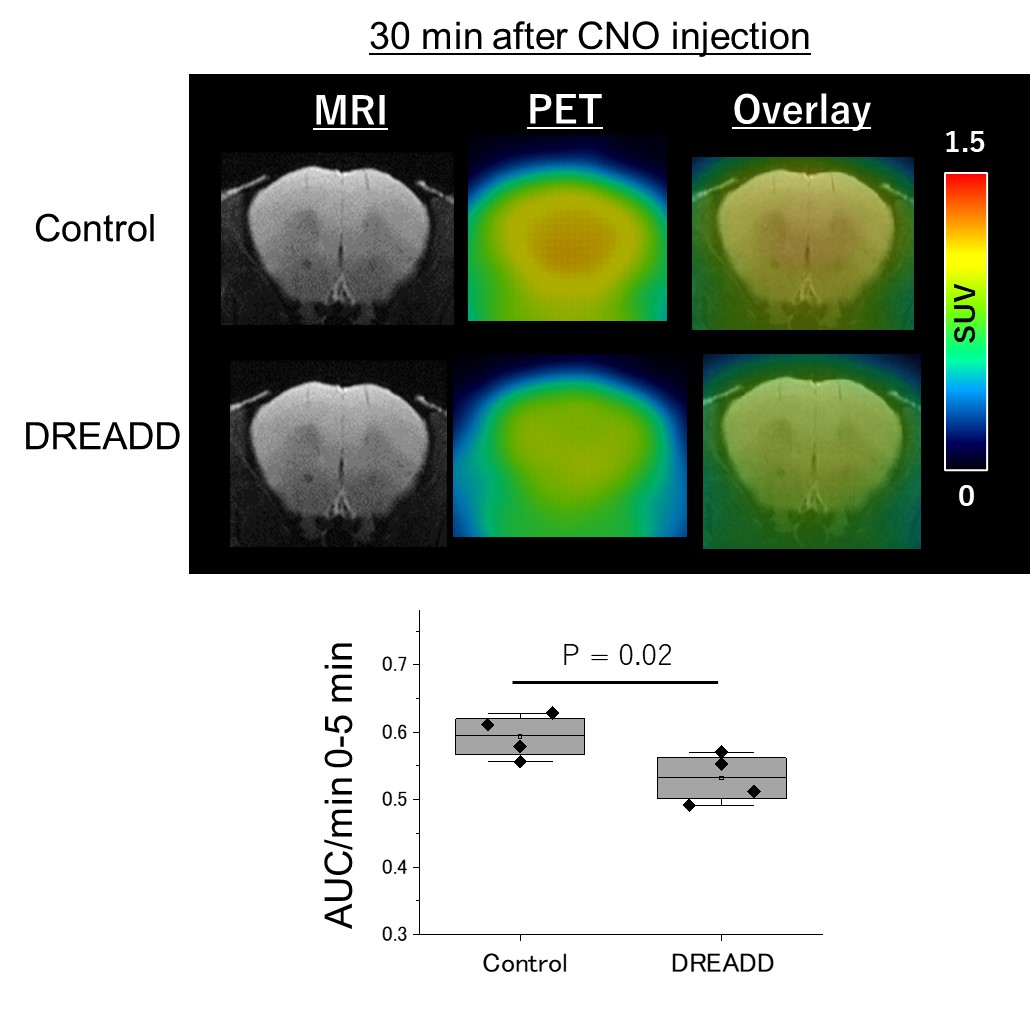
Animals:All the mice received viruses of AAV2/5-gfaABC1D-cyto-GCaMP6f specifically in lateral orbitofrontal cortex (lOFC) astrocytes. Mice in DREADD group also received viruses of AAV2/5 GfaABC1D-hM3Dq-mCherry for in vivo Gq-GPCR/Ca2+ stimulation, , and control mice had only GCaMP viruses (Fig.1). 1 mg/kg body weight of clozapine N-oxide (CNO), an agonist of DREADDs, was administered by intraperitoneal injection. HP-MR:1-13C pyruvic acid was hyperpolarized using SpinAlingner following the standard protocol6. Anatomical MRI and 13C-chemical shift imaging were obtained using a Bruker 3T scanner using the homemade awake tool (Fig.1). We used the above-mentioned mice for HP-MR experiments. 30 minutes after the injection of CNO, we injected hyperpolarized 1-13C pyruvate at the awake condition via tail vein (Fig.2). Lactate to pyruvate and bicarbonate to pyruvate ratios were quantified and compared between the two groups. 18F-FDG-PET:At 30 min after the CNO administration, 18F-FDG (5–7 MBq/head, Fujifilm) was injected via the tail vein catheter, and simultaneously dynamic emission scans were performed for 60 min using a small-animal PET scanner (Inveon, Siemens Healthineers). Time-activity curves (TACs) of 18F-FDG were acquired using PMOD software (version 3.4, PMOD Technology) from the volumes of interest, which were manually described on the OFC. The radioactivity was decay-corrected to the injection time and is expressed as a standardized uptake value (SUV), normalized for injected radioactivity and body weight. The apparent initial uptake rate was calculated by dividing the area under the TAC (AUC) between 0 and 5 min by the minutes.RESULTS
HP-MR:Compared to the control group, the DREADD group showed decreased lactate/pyruvate and bicarbonate/pyruvate ratios (p < 0.05), indicating noticeable metabolic shifts upon DREADD-induced perturbations (Fig.2). 18F-FDG-PET: Apparent initial uptake rates, defined by AUC (0 to 5 min) per min, were decreased in the DREADD group when CNO was injected 30 min before the 18F-FDG injection (p = 0.02) (Fig.3).DISCUSSION
By elevating the Ca2+ of astrocytes in the specific brain regions through DREADD, a reduction was observed in the flux from HP-MR 13C pyruvate in the same area and in the 18F-FDG-PET signal, suggesting the possibility that fluctuations in astrocyte Gq-GPCR/ Ca2+ signaling may contribute to those imaging data. The contribution of astrocytes to the 18F-FDG-PET signal has been demonstrated in studies involving the administration of ceftriaxone4 and clozapine5. These studies have reported that changes in 18F-FDG accumulation occurred by altering Glutamate transporter 1 (GLT1), and it is being discussed that 18F-FDG signals do not solely originate from neuronal activity. Further studies are needed to determine how GLT1 and Monocarboxylate Transporter 1(MCT1, utilized for the uptake of pyruvate from the blood (Fig.4)) change when Ca2+ in astrocytes fluctuates, as shown in this study.CONCLUSION
We reported that fluctuation of astrocyte Ca2+ contributes to the metabolism of HP-MR 13C pyruvate in the brain. It might be essential to consider astrocyte signaling in interpreting HP-MR 13C pyruvate, as was shown in 18F-FDG-PET studies4,5.Acknowledgements
This work was supported by AMED under grant no. JP22dk0207063 and MEXT Quantum Leap Flagship Program (MEXT QLEAP) under grant no. JPMXS0120330644.References
1. Hirata et al. Ann of Neuro 2023 (online ahead of print) 2. Pellerin L, et al. Proc Natl Acad Sci U S A 1994; 91: 10625–10629 3. Roth. Neuron 2016;89(4):683-94 4. Zimmer et al. Nature Neurosci 2017; 20(3):393-395. 5. Rocha et al. Eur J Nucl Med Mol Imaging. 2022;49(7):2251-2264. 6. SpinAligner User Manual v5Figures