0162
Prolonged Central Thalamic Intermittent Theta-Burst Stimulation Rescued Memory Deficits in Alzheimer's Disease Mouse Model1Department of Biomedical Engineering, National Yang Ming Chiao Tung University, Taipei, Taiwan, 2PhD Program in Medical Neuroscience, College of Medical Science and Technology, Taipei Medical University, Taipei, Taiwan, 3School of Medicine, College of Medicine, National Yang Ming Chiao Tung University, Taipei, Taiwan, 4Abbott Neuromodulation, Austin, TX, United States, 5Department of Neurology, Hualien Tzu Chi Hospital, Buddhist Tzu Chi Medical Foundation, Hualien, Taiwan, 6Department of Neurology, School of Medicine, Tzu Chi University, Hualien, Taiwan
Synopsis
Keywords: Alzheimer's Disease, Alzheimer's Disease, Intermittent theta-burst stimulation (iTBS)
Motivation: Addressing the global AD crisis by investigating CT-iTBS as a non-pharmacological treatment to enhance memory and cognition.
Goal(s): To explore the therapeutic efficacy and determine the optimal treatment protocol of CT-iTBS in AD while unveiling its potential underlying mechanism for enhancing memory and cognitive functions.
Approach: Utilized brain magnetic resonance imaging analysis, behavioral tests, and immunofluorescence staining for assessing the therapeutic effect of different durations of CT-iTBS treatment.
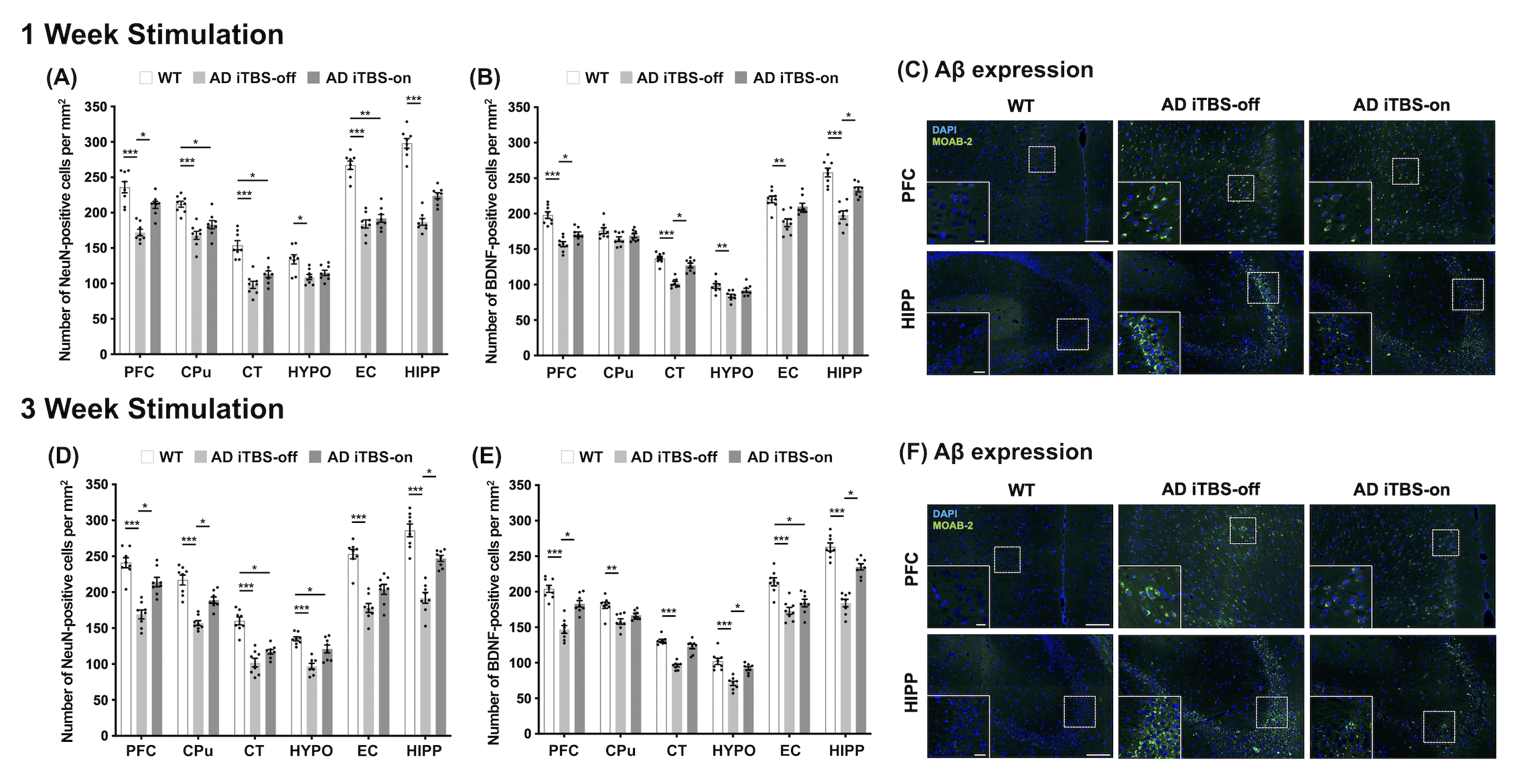
Results: Prolonged CT-iTBS significantly enhanced cognitive and memory behaviors, altered brain functional connectivity, promoted a neuroprotective effect, and reduced amyloid accumulation in AD mouse model. These findings present a promising therapeutic avenue for AD patients.
Impact: Our findings revealed a highly promising avenue for enhancing the quality of life for individuals with AD and provided insights into the potential underlying neuroprotective mechanisms of CT-iTBS in alleviating memory deficits.
Introduction
Alzheimer's disease (AD) has emerged as a global public health crisis over the past decades1,2, yet it continues to lack sufficient pharmacological treatments for potential cures3,4,5. In light of these constraints, some researchers are shifting their focus towards non-pharmacological therapy, deep brain stimulation (DBS)6,7. Previous research has provided compelling evidence of DBS's potential to enhance memory and cognitive function in AD through neuromodulation8,9,10 and neuroprotective effects11,12. By upregulating the expression of brain-derived neurotrophic factor (BDNF), which associated with neuronal growth, survival, and plasticity, DBS effectively enhance neuronal plasticity, providing evidence for mechanistic basis of its memory-enhancing effects13,14,15. Recently, a specific stimulation paradigm, intermittent theta-burst stimulation (iTBS), has been proposed as a more efficacious method for altering cognitive functions16,17. By delivering in rhythmic bouts of 3-8 Hz, iTBS recapitulates natural brain rhythm to immediately provoke theta oscillations in the brain, which have profound functional relevance to cognition18,19. However, the effectiveness of iTBS in ameliorating memory deficits in AD patients remains unknown. In this study, we targeted central thalamus (CT)20,21,22 as a promising site for DBS therapy in AD and explored two different stimulation durations to optimize treatment outcomes. We employed functional magnetic resonance imaging (fMRI) to evaluate the efficacy of iTBS in modifying brain activation. Subsequently, we validated the therapeutic potential of CT-iTBS in triple-transgenic AD model (3×TgAD) using behavioral tests, fMRI, and immunofluorescence (IF) staining to elucidate the treatment's effectiveness and underlying mechanism.Methods
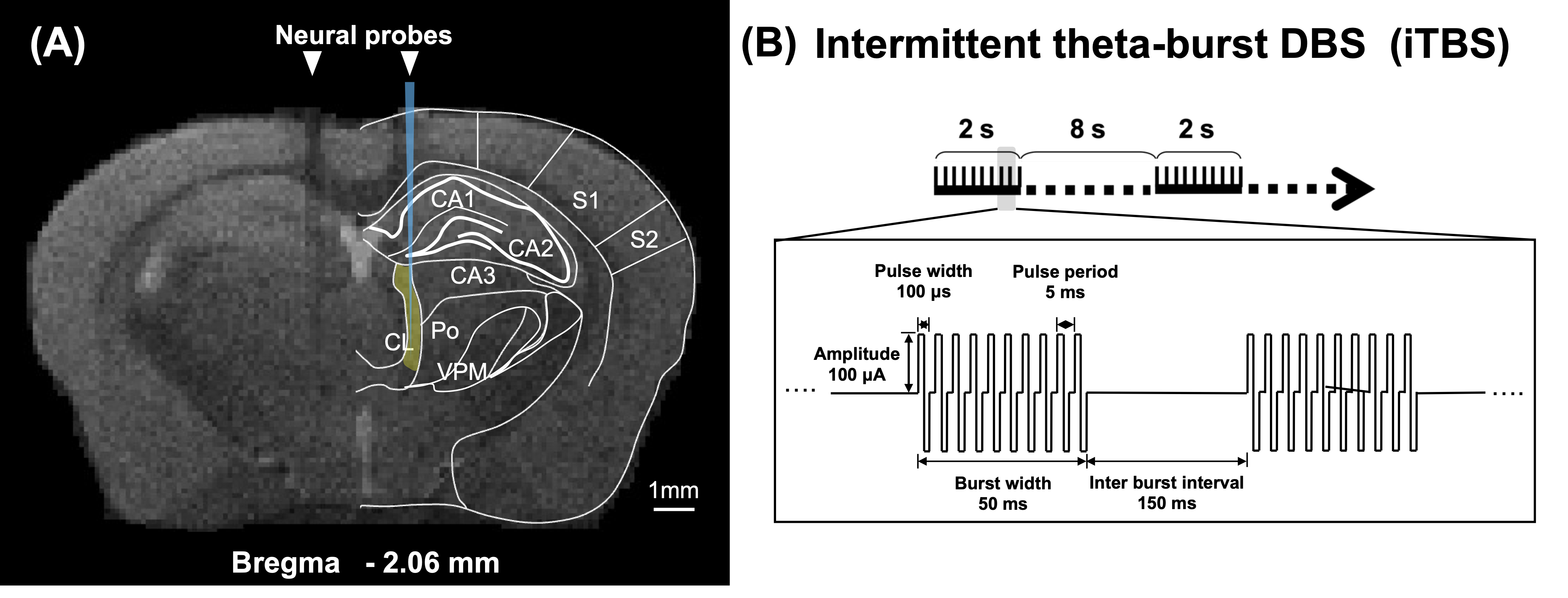
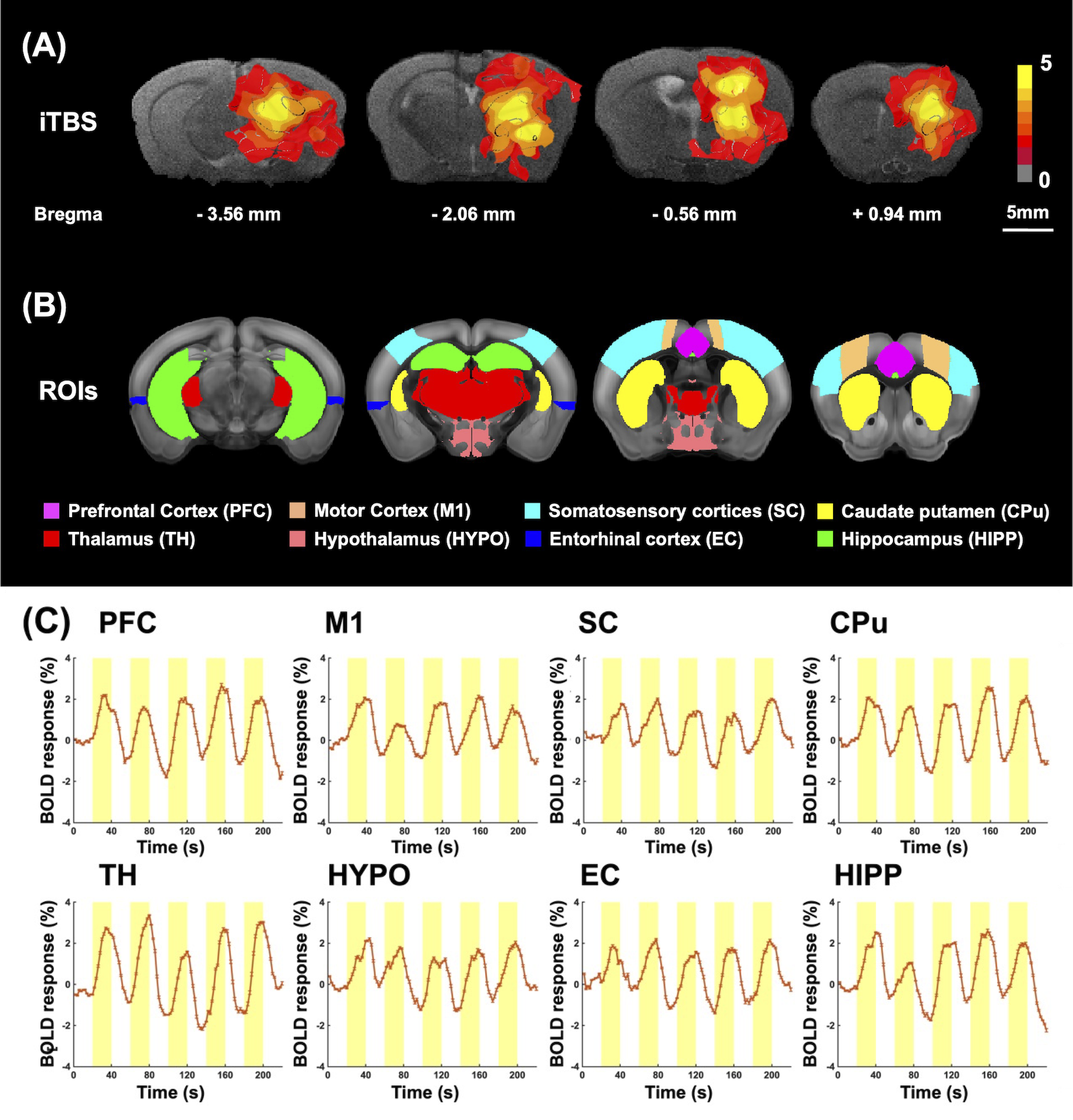
To identify the activated brain regions during unilateral CT-iTBS, we conducted MRI scanning on five adult C57BL/6 mice. For assessing the therapeutic effect of CT-iTBS, we used 8-month-old 3×TgAD mice as the AD disease model and age-matched wild-type mice as counterparts. Two stimulation protocols were implemented: one-week and three-week stimulation, each consisting of three groups—AD iTBS-off (N = 8), AD iTBS-on (N = 8), and wild-type (WT) (N = 8). All mice underwent implantation surgery with two MRI-compatible neural probes inserted into bilateral CT (AP: −1.56 mm, ML: ± 0.7 mm, DV: −3.0 mm). After a one-week recovery period, mice received one-week or three-week bilateral CT-iTBS for thirty minutes daily, with stimulation parameters as shown in Figure 1. Anxiety, long-term recognition memory, and working memory were assessed using the open field test (OFT), novel object recognition test (NOR), and T-maze test. MRI images were acquired using a 7 Tesla Bruker MRI system. fMRI data were collected through a gradient-echo planar imaging sequence with specific parameters: repetition time = 2,000 ms, echo time = 20 ms, 14 coronal slices, slice thickness = 0.5 mm, field of view = 20 × 20 mm2, matrix size = 80 × 80. Regions of interest (ROIs) included the prefrontal cortex (PFC), somatosensory cortices (SC), motor cortex (M1), caudate putamen (CPu), hippocampus (HIPP), hypothalamus (HYPO), thalamus (TH), and entorhinal cortex (EC). Functional connectivity (FC) was analyzed using the FMRIB Software Library v6.0 and the Analysis of Functional NeuroImages software. The average number of positive cells per region in IF staining was calculated using Fiji/ImageJ software. Statistical analysis was performed with Kruskal–Wallis tests and Dunn's multiple comparisons test, and significance was determined at a p-value < 0.05. Data were presented as mean ± standard error of the mean.Results
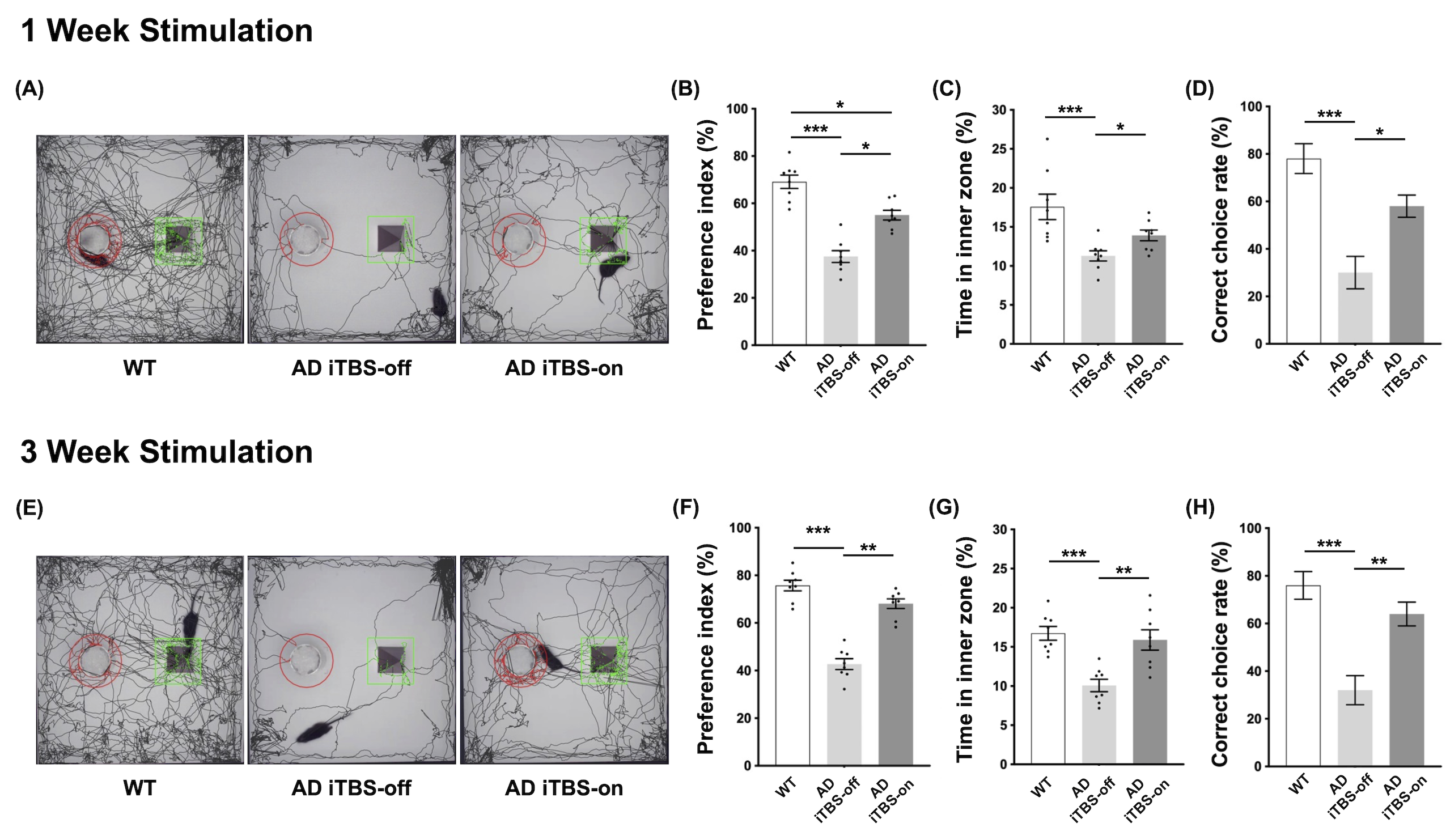
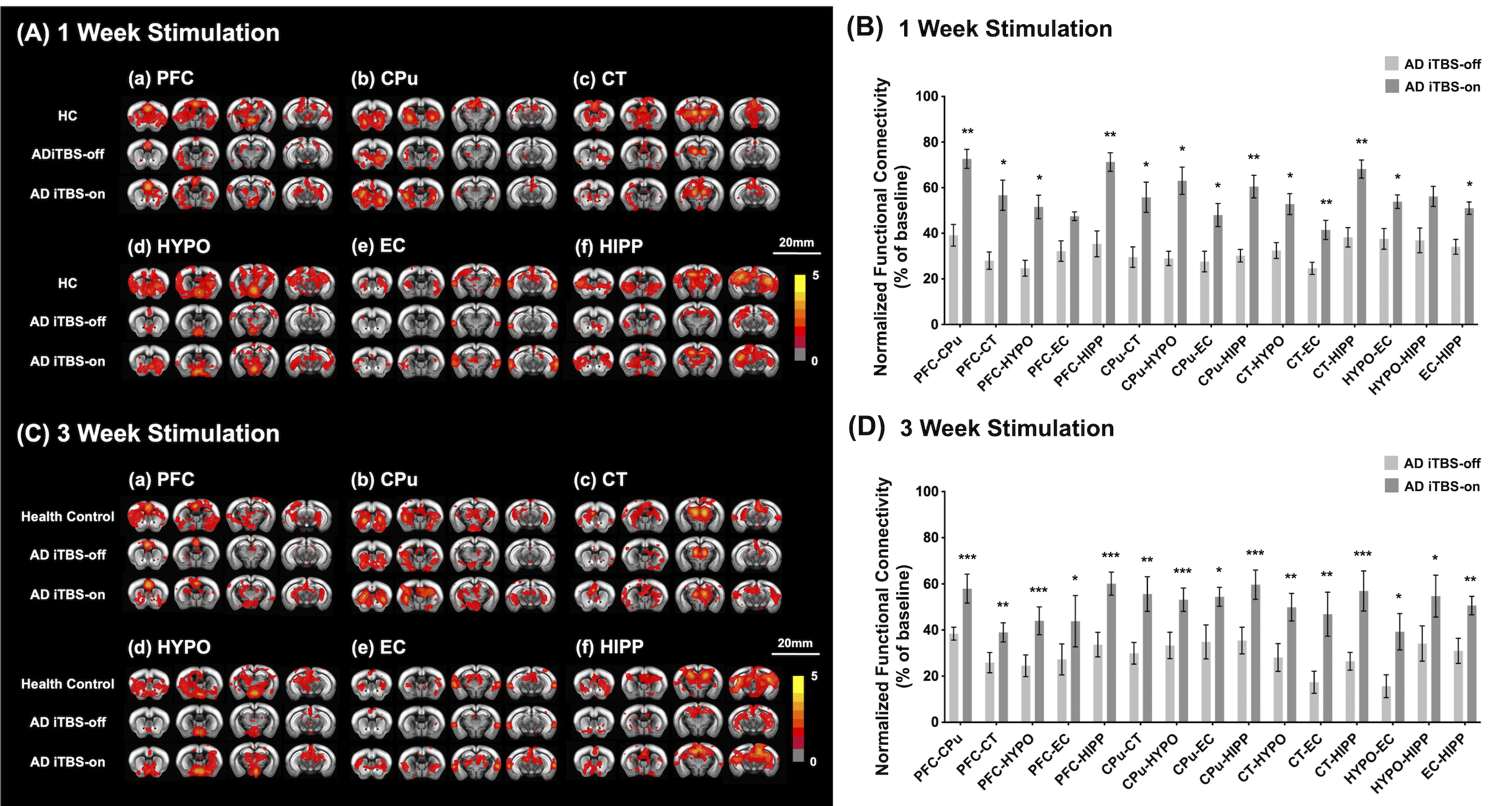
CT-iTBS activated a variety of cortical and subcortical regions, eliciting robust and well-synchronized responses in alignment with the stimulation paradigm (Figure 2). Applying CT-iTBS treatment to AD mouse resulted in significant improvements in memory and cognitive performance, particularly in AD iTBS-on groups, especially after three-week treatment (Figure 3). rsfMRI revealed significantly higher FC between PFC, HIPP, EC, and CPu following both durations of CT-iTBS treatment, indicating enhanced corticolimbic circuits contributing to improved memory cognitive function (Figure 4). IF staining unveiled an increase in neurotrophic factors and neuronal cell density following CT-iTBS, suggesting a neuroprotective effect. Additionally, the accumulation of beta-amyloid in PFC and HIPP significantly decreased after three-week of CT-iTBS (Figure 5).Discussion
The broad region activated by CT-iTBS at cortical (ACC and M1) and subcortical (CPu, Hip, CL, and HypoTH) were associated with corticolimbic, corticostriatal, and thalamocortical networks23,24,25, which were related to memory cognitive function. After applying CT-iTBS in AD mouse model, enhancement of BDNF expression in memory-related brain regions, fostering a neuroprotective effect and neuron plasticity26,27,28. These changes were supported by the alterations of FC within memory and cognitive networks, paralleling improved behavioral performances.Conclusion
Our study sheds light on the verification of efficacy and potential underlying mechanism of CT-iTBS. Additionally, our findings indicate that prolonging CT-iTBS resulted in more prominent therapeutic outcomes, highlighting its potential to alleviate symptoms in individuals with AD.Acknowledgements
This work is financially supported by National Science and Technology Council under Contract numbers of NSTC 112-2622-8-A49 -010 -TE2, 111-2221-E-A49 -049 -MY2, 112-2314-B-303 -016 -, 112-2321-B-A49 -009 -.References
1. Aranda MP, Kremer IN, Hinton L, et al. Impact of dementia: Health disparities, population trends, care interventions, and economic costs. Journal of the American Geriatrics Society. 2021;69(7):1774-1783.
2. Livingston G, Huntley J, Sommerlad A, et al. Dementia prevention, intervention, and care: 2020 report of the Lancet Commission. The Lancet. 2020;396(10248):413-446.
3. Zvěřová M. Clinical aspects of Alzheimer's disease. Clinical biochemistry. 2019;72:3-6.
4. Cummings J, Lee G, Nahed P, et al. Alzheimer's disease drug development pipeline: 2022. Alzheimer's & Dementia: Translational Research & Clinical Interventions. 2022;8(1):e12295.
5. Asher S, Priefer R. Alzheimer's disease failed clinical trials. Life Sciences. 2022:120861.
6. Lyketsos CG, Targum SD, Pendergrass JC, Lozano AM. Deep brain stimulation: a novel strategy for treating Alzheimer’s disease. Innovations in clinical neuroscience. 2012;9(11-12):10.
7. Yu D, Yan H, Zhou J, Yang X, Lu Y, Han Y. A circuit view of deep brain stimulation in Alzheimer’s disease and the possible mechanisms. Molecular Neurodegeneration. 2019;14:1-12.
8. Remoli G, Tariciotti L, Remore LG, et al. An updated overview of recent and ongoing deep brain stimulation (DBS) trials in patients with dementia: a systematic review. Neurological Sciences. 2023:1-33.
9. Pople CB, Meng Y, Li DZ, et al. Neuromodulation in the treatment of Alzheimer’s disease: Current and emerging approaches. Journal of Alzheimer's Disease. 2020;78(4):1299-1313.
10. Luo Y, Sun Y, Tian X, et al. Deep brain stimulation for Alzheimer's disease: stimulation parameters and potential mechanisms of action. Frontiers in Aging Neuroscience. 2021;13:619543.
11. McKinnon C, Gros P, Lee DJ, et al. Deep brain stimulation: potential for neuroprotection. Annals of clinical and translational neurology. 2019;6(1):174-185.
12. Park H, Poo M-m. Neurotrophin regulation of neural circuit development and function. Nature Reviews Neuroscience. 2013;14(1):7-23.
13. Sun Z, Jia L, Shi D, et al. Deep brain stimulation improved depressive-like behaviors and hippocampal synapse deficits by activating the BDNF/mTOR signaling pathway. Behavioural Brain Research. 2022;419:113709.
14. Pohodich AE, Yalamanchili H, Raman AT, et al. Forniceal deep brain stimulation induces gene expression and splicing changes that promote neurogenesis and plasticity. Elife. 2018;7:e34031.
15. Eyileten C, Sharif L, Wicik Z, et al. The relation of the brain-derived neurotrophic factor with microRNAs in neurodegenerative diseases and ischemic stroke. Molecular neurobiology. 2021;58(1):329-347.
16. Suppa A, Huang Y-Z, Funke K, et al. Ten years of theta burst stimulation in humans: established knowledge, unknowns and prospects. Brain stimulation. 2016;9(3):323-335.
17. Sweet JA, Eakin KC, Munyon CN, Miller JP. Improved learning and memory with theta‐burst stimulation of the fornix in rat model of traumatic brain injury. Hippocampus. 2014;24(12):1592-1600.
18. Shirvalkar PR, Rapp PR, Shapiro ML. Bidirectional changes to hippocampal theta–gamma comodulation predict memory for recent spatial episodes. Proceedings of the National Academy of Sciences. 2010;107(15):7054-7059.
19. Miller JP, Sweet JA, Bailey CM, Munyon CN, Luders HO, Fastenau PS. Visual-spatial memory may be enhanced with theta burst deep brain stimulation of the fornix: a preliminary investigation with four cases. Brain. 2015;138(7):1833-1842.
20. Sieveritz B, Raghavan RT. The Central Thalamus: Gatekeeper or Processing Hub? Journal of Neuroscience. 2021;41(23):4954-4956.
21. Lopez J, Wolff M, Lecourtier L, et al. The intralaminar thalamic nuclei contribute to remote spatial memory. Journal of Neuroscience. 2009;29(10):3302-3306.
22. Arnts H, Coolen SE, Fernandes FW, et al. The intralaminar thalamus: a review of its role as a target in functional neurosurgery. Brain Communications. 2023:fcad003.
23. Simioni AC, Dagher A, Fellows LK. Effects of levodopa on corticostriatal circuits supporting working memory in Parkinson's disease. Cortex. 2017;93:193-205.
24. Quidé Y, Morris RW, Shepherd AM, Rowland JE, Green MJ. Task-related fronto-striatal functional connectivity during working memory performance in schizophrenia. Schizophrenia research. 2013;150(2-3):468-475.
25. Staudigl T, Zaehle T, Voges J, et al. Memory signals from the thalamus: early thalamocortical phase synchronization entrains gamma oscillations during long-term memory retrieval. Neuropsychologia. 2012;50(14):3519-3527.
26. Bramham CR, Messaoudi E. BDNF function in adult synaptic plasticity: the synaptic consolidation hypothesis. Progress in neurobiology. 2005;76(2):99-125.
27. Lu B, Nagappan G, Lu Y. BDNF and synaptic plasticity, cognitive function, and dysfunction. Neurotrophic factors. 2014:223-250.
28. Kowiański P, Lietzau G, Czuba E, Waśkow M, Steliga A, Moryś J. BDNF: a key factor with multipotent impact on brain signaling and synaptic plasticity. Cellular and molecular neurobiology. 2018;38:579-593.
Figures