0132
A different interpretation of the DIANA fMRI signal1Massachusetts Institute of Technology, Cambridge, MA, United States
Synopsis
Keywords: Probes & Targets, Brain Connectivity
Motivation: A recent study argued that it is possible to detect neuroelectrical potentials using an fMRI scanning approach called DIANA. Although DIANA signals coincide with electrophysiological measurements, no mechanism for the effect was reported
Goal(s): We sought to implement DIANA in order to understand the origins of the reported results.
Approach: We applied variants of the DIANA pulse sequence under test and control conditions, comparing results with simulations.
Results: We observed DIANA signals but show that neural activity is neither necessary nor sufficient for this. Instead, the DIANA signal appears to result largely from nonideal aspects of pulse sequence timing.
Impact: Our study suggests that the DIANA signal arises from an artifact in the line scan pulse sequence. This indicates possible pitfalls in implementing such sequence designs and emphasizes the continuing need for fMRI-based direct readouts of neural activity.
Introduction
A recent study (Ref1) argued that it is possible to detect neuroelectrical potentials using a specialized fMRI scanning approach called DIANA. We want to implement DIANA in anesthetized rats and measure responses to somatosensory stimulation to reproduce core findings of the original study in order to understand the mechanism of the signal.Method
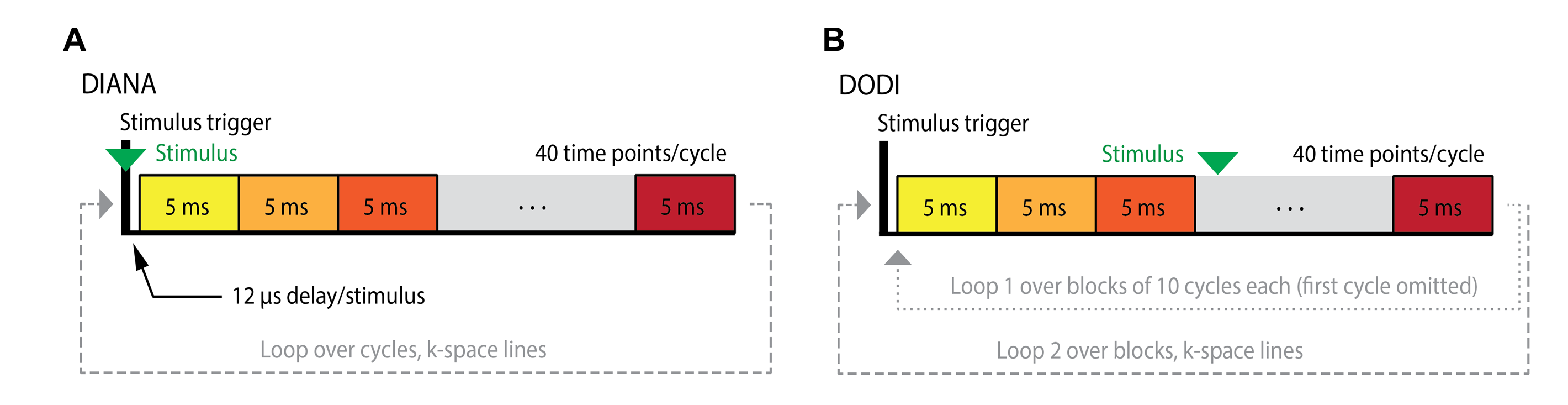
We noticed that the DIANA sequence used by Toi et al. includes a short time interval of 12 µs associated with triggering that is only present in line scans synchronized with stimulation, once every 200 ms. To test whether omission of the triggering interval changes the DIANA contrast, we designed an alternative pulse sequence which we call “DIANA omitting disruptive intervals” (DODI) which does not have a TR delay of additional 12 µs. Hind paw stimulation in rats was test under various conditions: (1) with stimulation; (2) under non-stimulated conditions by omitting the trigger in the pulse sequence; (3) where the stimulation trigger was retained but the stimulation cable was unplugged from the stimulation amplifier, (4) postmortem and (5) 1h postmortem. DIANA fMRI experiments were implemented using the exact pulse sequence (ppg file) utilized by Toi et al. or closely modified from the standard Bruker FLASH ppg file to confirm the deterministic importance of the stimulus trigger delay in giving rise to the DIANA signal. Figure 1 shows the implementation both DIANA and DODI sequence. A solenoid coil with a diameter of 1 cm was placed inside the Falcon tube and connected to the same stimulator used for functional imaging experiments in rats. Imaging was then performed in the presence of stimulation with parameters identical to those used in DIANA and DODI experiments to test for synchrony between magnetic induction caused by the coil and stimulation regime for confirmation of 5ms time resolution of both sequences. MRI signal from DIANA and DODI acquisition schemes was simulated by integrating the Bloch equations (Ref2) in a rotating frame demodulated by the resonance frequency, using a custom script coded in Python.Results
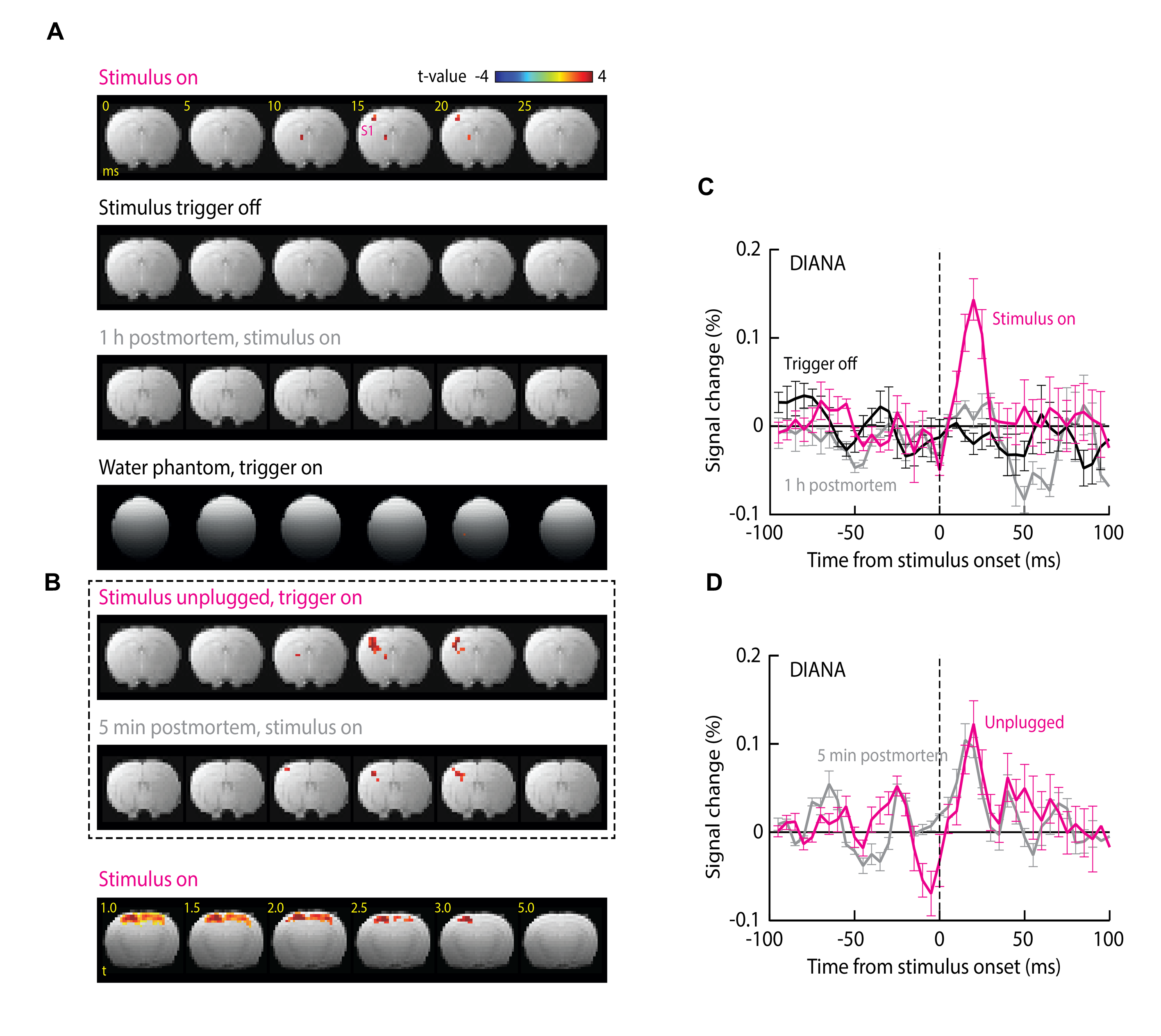
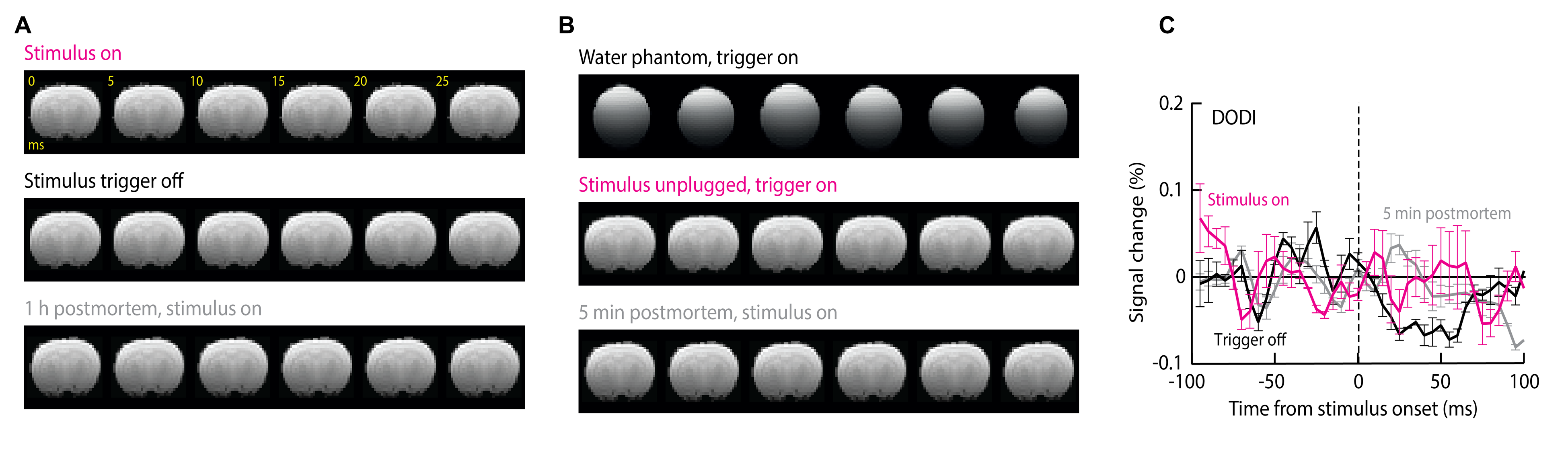
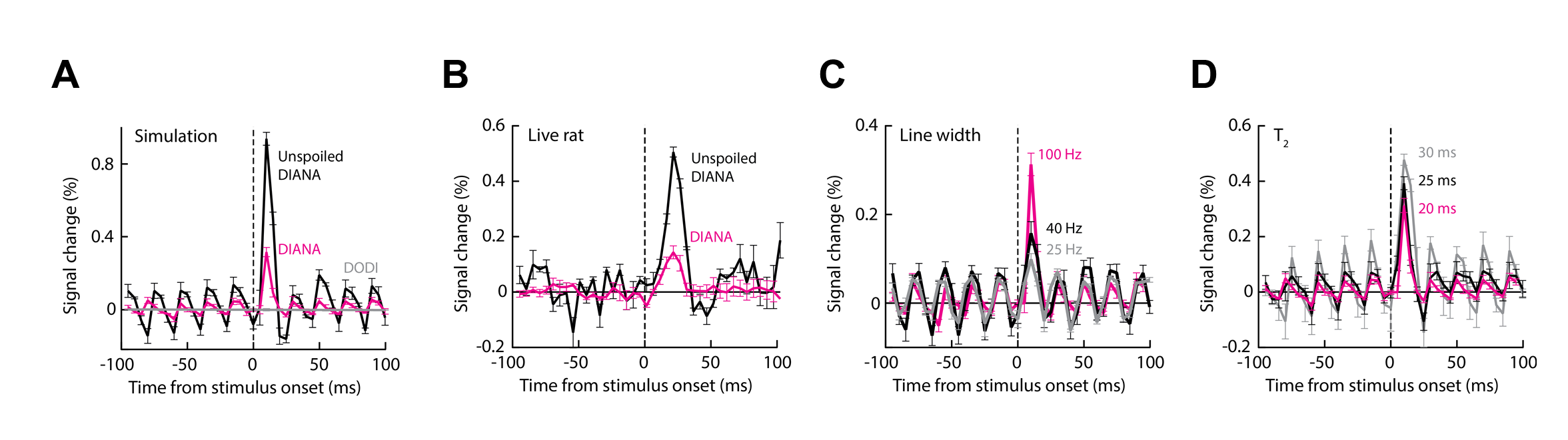
We found that DIANA produced reproducible, transient responses in primary somatosensory cortex (S1) that follow the stimulus with an average time-to-peak of 18.0±1.3 ms and amplitude of 0.14±0.02% (Fig. 2 A,B); the mean response was statistically significant with t-test p = 0.002 (n=5). At a t-value threshold of 1.0, the DIANA-associated signal changes appeared to stretch across most of the dorsal cortex, but at a more conservative t threshold of 3.0, the signal was restricted to a contralateral S1 region where peak activity is expected (5) (Fig. 1C). No DIANA-associated responses (t-test p ≥ 0.12, n=5) were seen in control experiments performed without stimulus triggering, in animals imaged 1 h after death, or in a water phantom (Fig. 1D,E). Unexpectedly, however, significant responses of average magnitude 0.12 ± 0.03% (t-test p=0.007, n=5) were also observed in animals that underwent DIANA imaging with stimulus triggering but with the electrical stimulus cable unplugged and stimulation therefore disabled. A mean response of 0.10±0.02% (p=0.003, n=5) was seen furthermore in animals scanned five minutes after death, as defined by loss of periodic pulse oximetry signals. The experiments of Fig. 1B were repeated using the DODI approach, and resulted in no detectable fMRI signal changes (t-test p ≥ 0.16, n = 5) under any conditions (Fig. 2F-H). The simulation confirmed that small signal peaks could be observed within milliseconds of the DIANA stimulation trigger, but that analogous signals are absent from the simulated DODI signal (Fig. 2D). The magnitude but not the timing of the simulated DIANA signals is strongly affected by radiofrequency spoiling and off-resonance frequencies.Discussion
These observations (Fig. 2A,D) imply that neuronal activity is not necessary for producing MRI signal changes in the DIANA experiments in respect to the results in condition of unplugged stimulation cable and postmortem animal. Because these experiments use stimuli that are well known to evoke reliable neural responses in somatosensory cortex, the results of the DODI measurement thus indicate that neural activity is insufficient to produce DIANA-like fMRI responses under conditions where the short stimulus-triggering delay has been removed from the MRI pulse sequence. The results from both DIANA and DODI experiments in conjunction with the simulation results strongly implicate that DIANA signal stems from the disturbed steady state by the trigger interval delay at stimulation and causing coherent echoes.Conclusion
We show that neural activity is neither sufficient nor necessary to produce DIANA signal. We use a combination of control conditions and simulations to demonstrate that DIANA signals can arise from nonideal aspects of the pulse sequence.Acknowledgements
No acknowledgement found.References
Ref 1. P. T. Toi et al., In vivo direct imaging of neuronal activity at high temporospatial resolution. Science 378, 160-168 (2022)
Ref 2. F. Bloch, Nuclear induction. Phys Rev 70, 4604-4673 (1946)
Figures