0115
Distinct longitudinal brain white matter microstructure changes and associated polygenic psychiatric and neurodegenerative disorder risk1Department of Health and Functioning, Western Norway University of Applied Sciences, Bergen, Norway, 2NORMENT Centre for Psychosis Research, Division of Mental Health and Addiction, University of Oslo and Oslo University Hospital, Oslo, Norway, 3Mohn Medical Imaging and Visualization Centre (MMIV), Bergen, Norway, 4Faculty of Health, Medicine and Life Sciences, Maastricht University, Maastricht, Netherlands, 5Department of Psychiatric Research, Diakonhjemmet Hospital, Oslo, Norway, 6Department of Psychology, University of Oslo, Oslo, Norway, 7Department of Radiology, Haukeland University Hospital, Bergen, Norway, 8Department of Biomedicine, University of Bergen, Bergen, Norway, 9KG Jebsen Centre for Neurodevelopmental Disorders, University of Oslo, Oslo, Norway
Synopsis
Keywords: DWI/DTI/DKI, Brain, Ageing | White Matter | Microstructure | Brain Ageing | Polygenic Risk | Magnetic Resonance Imaging | Diffusion MRI
Motivation: White matter microstructural (WMM) changes are a crucial feature of ageing and disease development. There is yet no comprehensive mapping of such changes.
Goal(s): Providing an overview of WMM changes at different spatial scales, and relationship of these changes to polygenic risk scores (PGRS) of developing psychiatric disorders and Alzheimer's disease.
Approach: WMM metrics were estimated using multiple diffusion approaches, associated with age and PGRS, and ageing changes (inter-scan interval:2.44±0.73 years) assessed at different spatial scales.
Results: We find spatially distributed WMM-changes and PGRS-associations across the brain (most age-sensitive: central and cerebellar WMM). Brain longitudinal changes reflected disorder PGRS better than cross-sectional measures.
Impact: The manuscript details for the first time longitudinal WMM changes in a large longitudinal sample (UK Biobank, N=2,676), and provides the currently most comprehensive overview of PGRS associations with WMM change and WMM (using an additional cross-sectional validation sample, N=31,056).
Background
While there are multiple studies examining white matter microstructure in cross-sectional data1-2, well-powered studies examining white matter ageing are still lacking. Additionally, the different studies usually analyse either region-averaged or voxel-level changes, supplemented by global average scores. Studies giving a systematic overview across these levels of analysis, or beyond diffusion tensor imaging are still lacking3. Finally, while the field of imaging genetics is experiencing an immense growth, polygenic risk associations in longitudinal data are also a rarity. Thus, associating the polygenic risk of a given person with the trajectories of their brain changes is a valuable step of adding a practical component to identified patterns of changes. Additionally, at this point there is no studies giving a broad overview of white matter associations with polygenic risk scores of common brain and psychiatric disorders.Methods
We use the UK Biobank4 repeated measures diffusion MRI data of N = 2,678 participants (53.36% females), which was taken at two time-points with a mean inter-scan interval of 2.43±0.73 years. Additionally, we use the portion of the UK Biobank cross-sectional sample which was not included in the longitudinal sampling N = 31,056 as a validation sample for cross-sectional investigations of polygenic risk score associations with white matter metrics. After quality control exclusions, tract-based spatial statistics (TBSS)5 were estimated for conventional and advanced diffusion models, and statistics run within the confines of the fractional anisotropy white matter skeleton. The used biophysical models for white matter microstructure metric estimation included Diffusion Tensor Imaging6, Diffusion Kurtosis Imaging7, the Spherical Mean Technique8 and its multi-compartment extension9, the Bayesian Rotationally Invariant Approach10, and White Matter Tract Imaging11. We estimated polygenic risk scores for Alzheimer's disease and common psychiatric disorders, including Major Depressive Disorder, Bipolar Disorder, Anxiety Disorder, Autism Spectrum Disorder, Schizophrenia, Attention Deficit Hyperactivity Disorder, and Obsessive-Compulsive Disorder. These polygenic scores were informed by previous large-scale genome-wide association studies.Results
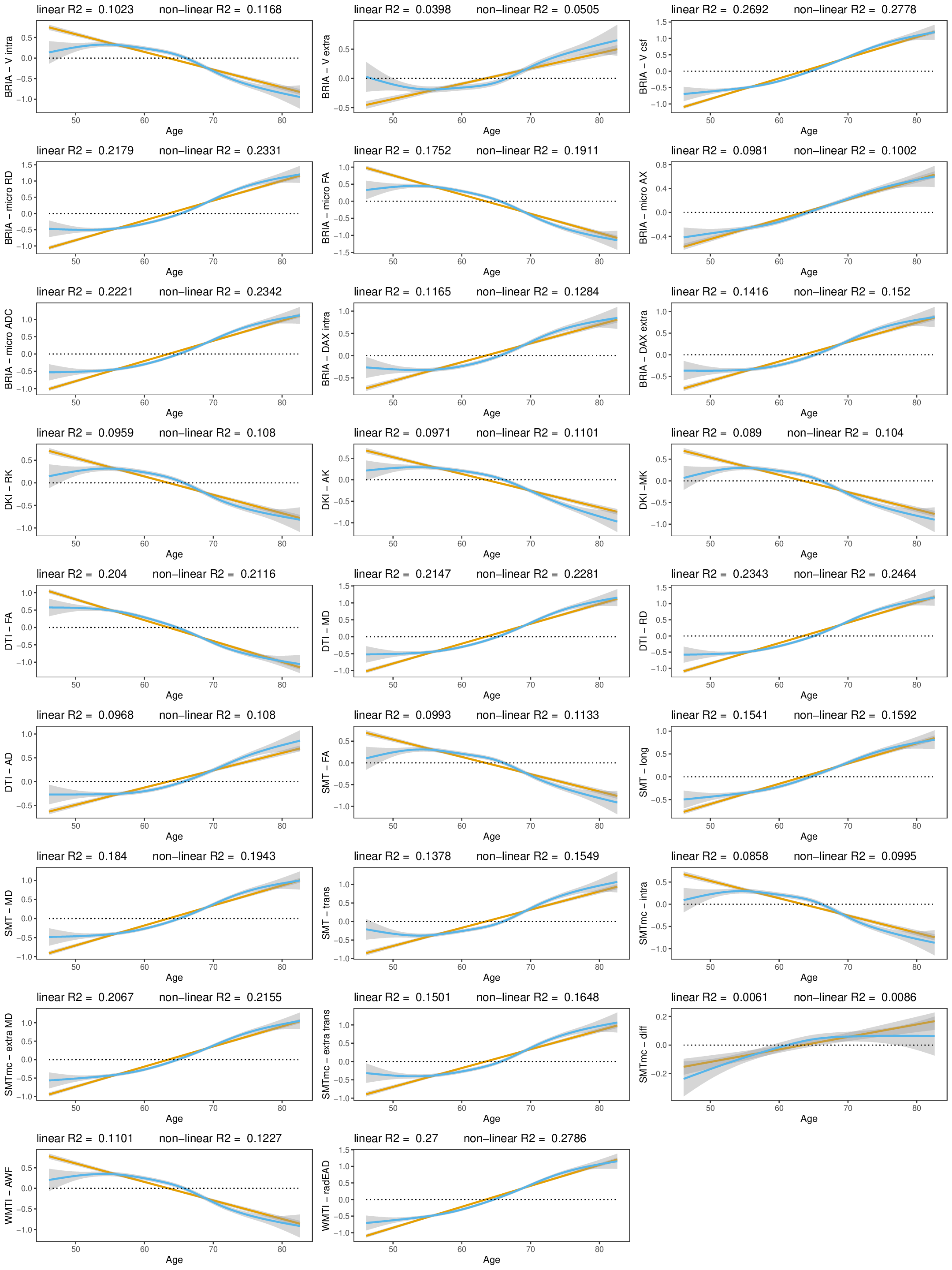
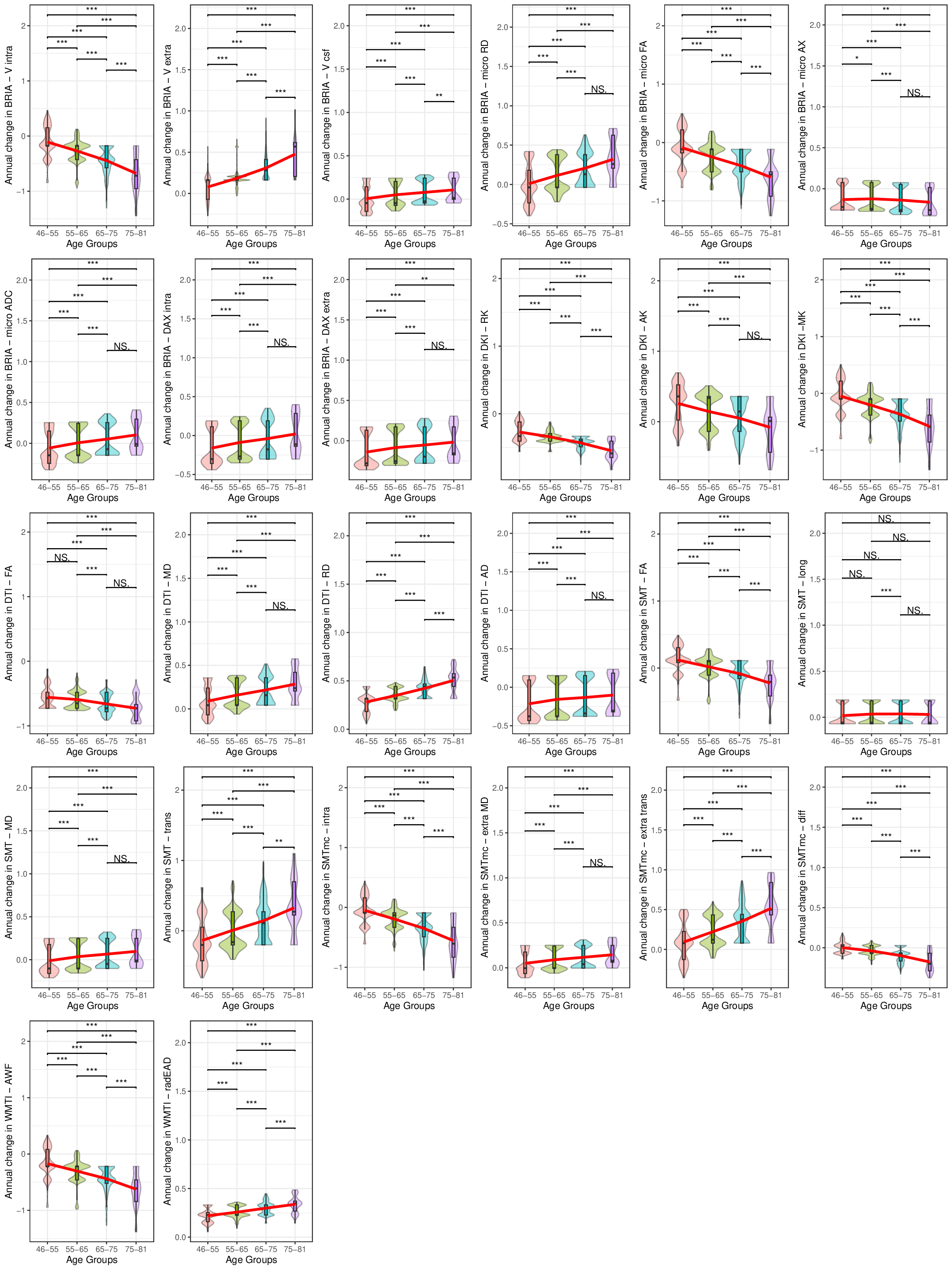
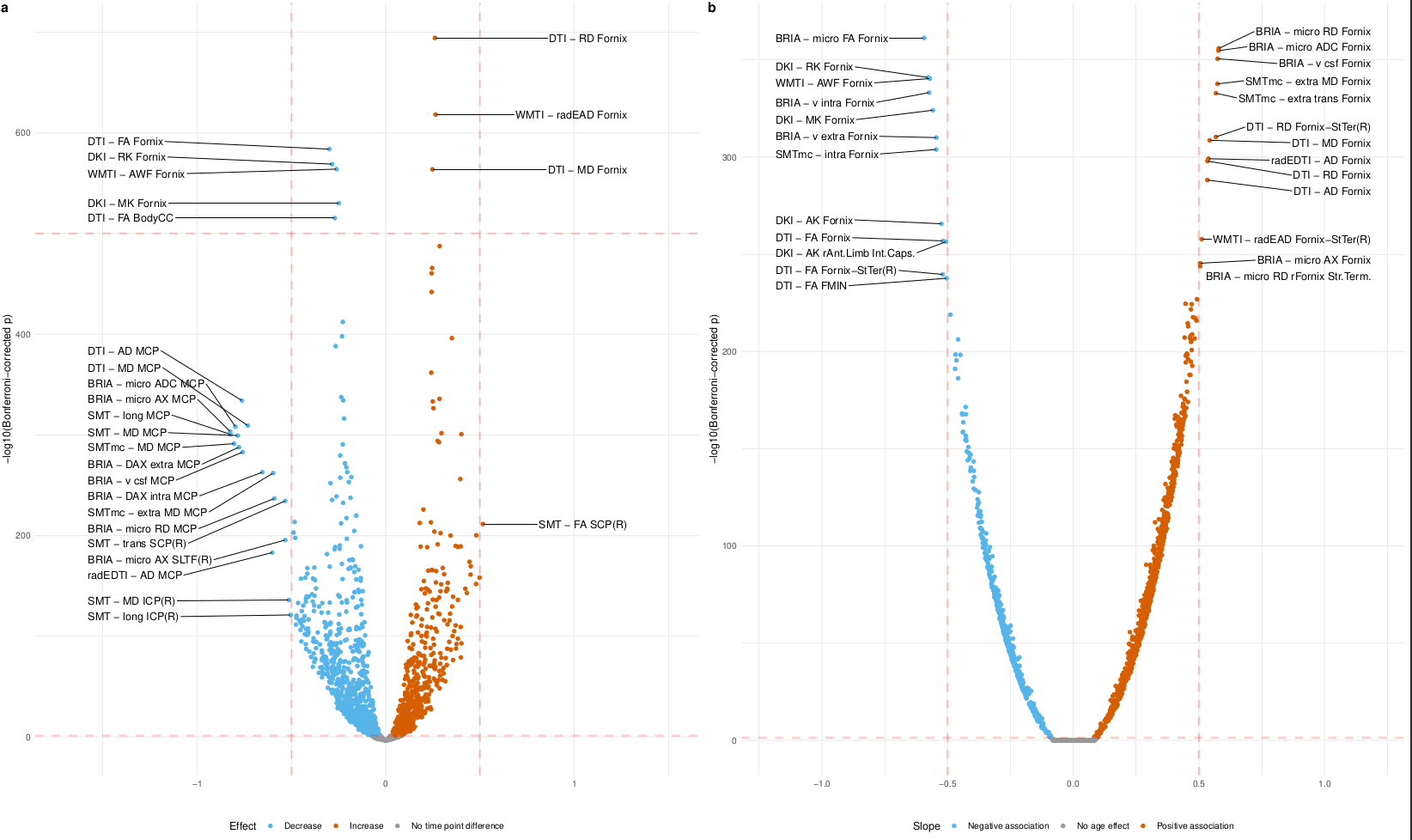
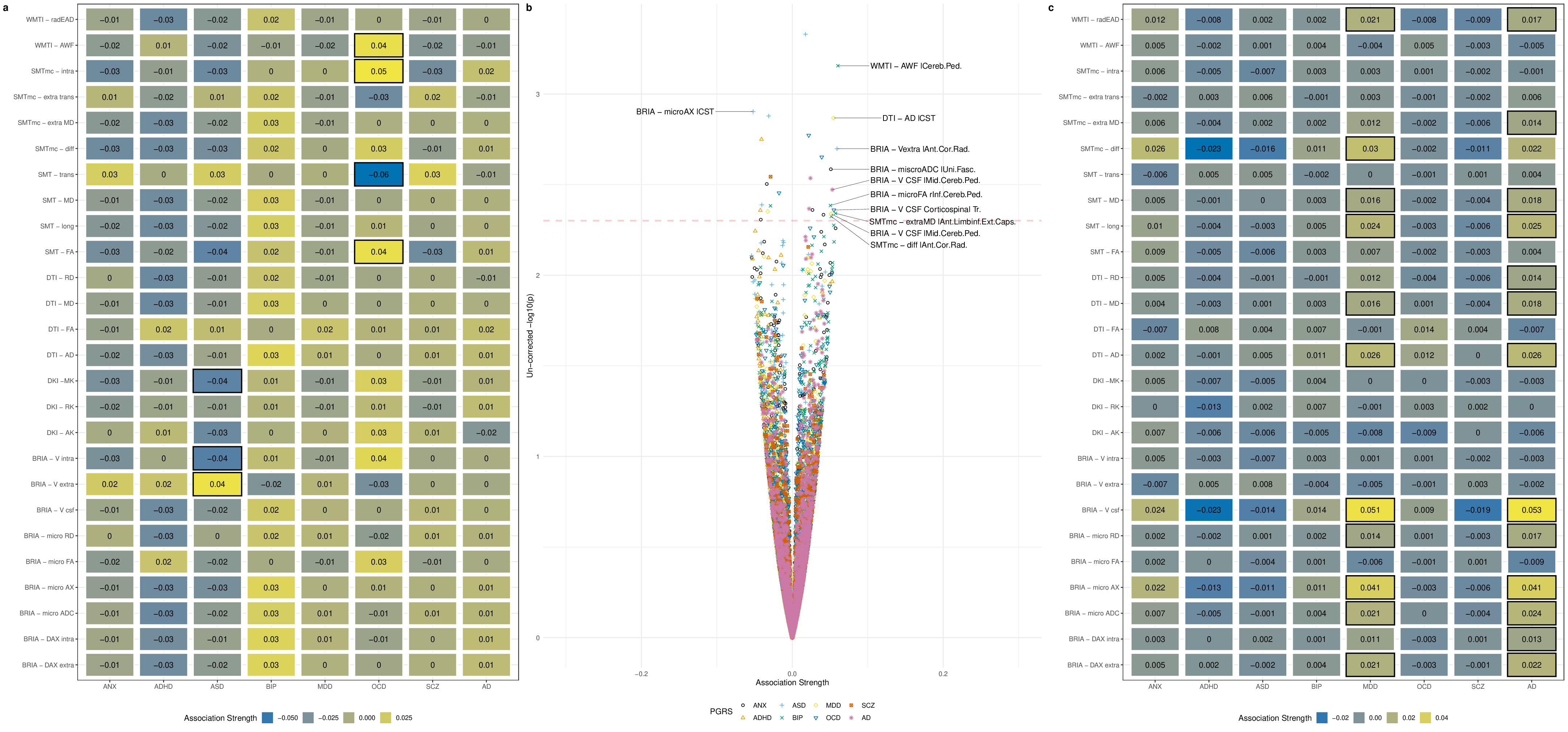
Our findings demonstrated lower global and regional fractional anisotropy, the intra-axonal water fraction, and kurtosis metrics at higher ages (Fig.1&3). In contrast, axial, radial, and mean diffusivity metrics as well as free water and extra-axonal water fractions appeared higher at higher age. These findings highlighted fornix and cingulate which presented strongest age-associations over time. Additionally, the annual rate of white matter change accelerated at higher ages for most examined diffusion metrics, indicating naturally occuring accelerated ageing at higher ages (Fig.2). Voxel-level analyses showed general trends of decreasing anisotropy, and variable spatial patterns for other diffusion metrics, indicating differential changes in frontal compared to other brain regions (Fig.4). By assessing associations of global and regional white matter change with polygenic risk scores, we provided practical implications of the presented age-related changes (Fig.5). We identified strongest and most consistently associated with Alzheimer's Disease polygenic risk scores with the annual change in inferior cerebral peduncle. Fornix and cingulate annual change presented some of the strongest polygenic risk associations with different psychiatric disorders.Conversely, considering cross-sectional measures of white matter, global averages of multiple diffusion metrics were particularly associated with Alzheimer's polygenic risk in the observed longitudinal sample, which was however not the case in the validation sample, which showed stronger associations with autism spectrum disorder. Regional cross-sectional white matter metrics showed spatially distributed associations with the polygenic risk of several disorders, consistently presenting strongest and most significant associations in the limbic system and brain stem. Finally and importantly, polygenic risk scores for Alzheimer's Disease and psychiatric disorders were stronger related to the annual rate of change than cross-sectional WM measures.Conclusion
The presented results indicate that demyelination processes and WM disintegration are heterogeneously distributed across the brain during midlife and older ages. Differences in water compartments' diffusivity suggest weaker plasticity in the frontal regions during this short time. Importantly, longitudinal changes of white matter reflected the genetic risk for disorder development better than the utilized cross-sectional measures. Moreover, the spatial pattern of these brain-gene associations is distributed, indicating that certain brain regions correspond more to one or a set of polygenic risk scores than others. For example, such as the Our results underline the importance of longitudinal data analyses in understanding of the ageing brain and its' genetic underpinnings.Acknowledgements
This study has been conducted using UKB data under Application 27412. UKB has received ethics approval from the National Health Service National Research Ethics Service (ref 11/NW/0382). The work was performed on the Service for Sensitive Data (TSD) platform, owned by the University of Oslo, operated and developed by the TSD service group at the University of Oslo IT-Department (USIT). Computations were performed using resources provided by UNINETT Sigma2 – the National Infrastructure for High Performance Computing and Data Storage in Norway. Finally, we want to thank all UKB participants and facilitators who made this research possible. \\This research was funded by the Research Council of Norway (#223273, #300767); the South-Eastern Norway Regional Health Authority (#2022080, #2019101); and the European Union's Horizon2020 Research and Innovation Programme (#847776, #802998).References
1. Korbmacher, M., de Lange, A. M., van der Meer, D., Beck, D., Eikefjord, E., Lundervold, A., ... & Maximov, I. I. (2023). Brain‐wide associations between white matter and age highlight the role of fornix microstructure in brain ageing. Human brain mapping. doi: https://doi.org/10.1002/hbm.26333
2. Korbmacher, M., van der Meer, D., Beck, D., de Lange, A. M. G., Eikefjord, E., Lundervold, A., ... & Maximov, I. I. (2023). Brain asymmetries from midlife to old adulthood and hemispheric brain age. bioRxiv, 2023-08. doi: https://doi.org/10.1101/2023.08.21.554103
3. Korbmacher, M., van der Meer, D., Beck, D., Askeland-Gjerde, D.E.., Eikefjord, E., Lundervold, A., ... & Maximov, I. I. (2023). Distinct longitudinal brain white matter microstructure changes and associated polygenic risk of common psychiatric disorders and Alzheimer's disease in the UK Biobank. medRxiv, 2023-10. doi: https://doi.org/10.1101/2023.10.19.23297257
4. Matthews, P. M., & Sudlow, C. (2015). The UK biobank. Brain, 138(12), 3463-3465. https://doi.org/10.1093/brain/awv335
5. Smith, S. M., Jenkinson, M., Johansen-Berg, H., Rueckert, D., Nichols, T. E., Mackay, C. E., ... & Behrens, T. E. (2006). Tract-based spatial statistics: voxelwise analysis of multi-subject diffusion data. Neuroimage, 31(4), 1487-1505. https://doi.org/10.1016/j.neuroimage.2006.02.024
6. Basser, P. J., Mattiello, J., & LeBihan, D. (1994). MR diffusion tensor spectroscopy and imaging. Biophysical journal, 66(1), 259-267.
7. Jensen, J. H., Helpern, J. A., Ramani, A., Lu, H., & Kaczynski, K. (2005). Diffusional kurtosis imaging: the quantification of non‐gaussian water diffusion by means of magnetic resonance imaging. Magnetic Resonance in Medicine: An Official Journal of the International Society for Magnetic Resonance in Medicine, 53(6), 1432-1440. https://doi.org/10.1002/mrm.20508
8. Kaden, E., Kruggel, F., & Alexander, D. C. (2016). Quantitative mapping of the per‐axon diffusion coefficients in brain white matter. Magnetic resonance in medicine, 75(4), 1752-1763. https://doi.org/10.1002/mrm.25734
9. Kaden, E., Kelm, N. D., Carson, R. P., Does, M. D., & Alexander, D. C. (2016). Multi-compartment microscopic diffusion imaging. NeuroImage, 139, 346-359. https://doi.org/10.1016/j.neuroimage.2016.06.002
10. Reisert, M., Kellner, E., Dhital, B., Hennig, J., & Kiselev, V. G. (2017). Disentangling micro from mesostructure by diffusion MRI: a Bayesian approach. NeuroImage, 147, 964-975. https://doi.org/10.1016/j.neuroimage.2016.09.058
11.
Fieremans,
E., Jensen, J. H., & Helpern, J. A. (2011). White matter
characterization with diffusional kurtosis imaging. Neuroimage,
58(1), 177-188. https://doi.org/10.1016/j.neuroimage.2011.06.006
Figures