0031
Choroid plexus atrophy in premanifest synucleinopathy using 7 Tesla MRI: an underlooked mechanism for α-synuclein accumulation?1Brainstem Imaging Laboratory, Department of Radiology, Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital and Harvard Medical School, Charlestown, MA, United States, 2Department of Neurology and Movement Disorders, APHM, Hôpital Universitaire Timone, Marseilles, France, 3Multiscale Imaging and Integrative Biophysics Unit, LBN, National Institute on Aging, NIH, Baltimore, MD, United States, 4Department of Neurology, Medical University of Innsbruck, Innsbruck, Austria, 5Department of Neurology, Massachusetts General Hospital and Harvard Medical School, Boston, MA, United States, 6Division of Sleep Medicine, Harvard University, Boston, MA, United States, 7Escuela Nacional de Estudios Superiores Unidad Juriquilla, Universidad Nacional Autónoma de México, Querétaro, Mexico
Synopsis
Keywords: Parkinson's Disease, High-Field MRI, Choroid Plexus, Synucleinopathy, RBD
Motivation: As opposed to Alzheimer's disease, the mechanism linking neurotoxic protein accumulation to alterations in neurofluid turnover and in neuroimmunity due to choroid plexus (ChP) changes is understudied in premanifest synucleinopathy.
Goal(s): To determine changes in the structure of ChP in premanifest synucleinopathy and to generate a ChP probabilistic atlas.
Approach: ChP in multi-contrast 7 Tesla images of 12 premanifest synucleinopathy and 12 sex-and-age-matched controls were evaluated in terms of volume and signal intensity.
Results: Reduced ChP volume in premanifest synucleinopathy suggested ChP atrophy that may result in neurofluid dynamics and neuroimmune function impairment; a probabilistic atlas of ChP was generated.
Impact: ChP atrophy observed in premanifest synucleinopathy using high-resolution multi-contrast 7-Tesla MRI suggests a potential role of ChP in pathophysiology of synucleinopathies, urging further investigation. Probabilistic ChP atlas may aid precise MRI localization in future studies of ChP in living humans.
Introduction
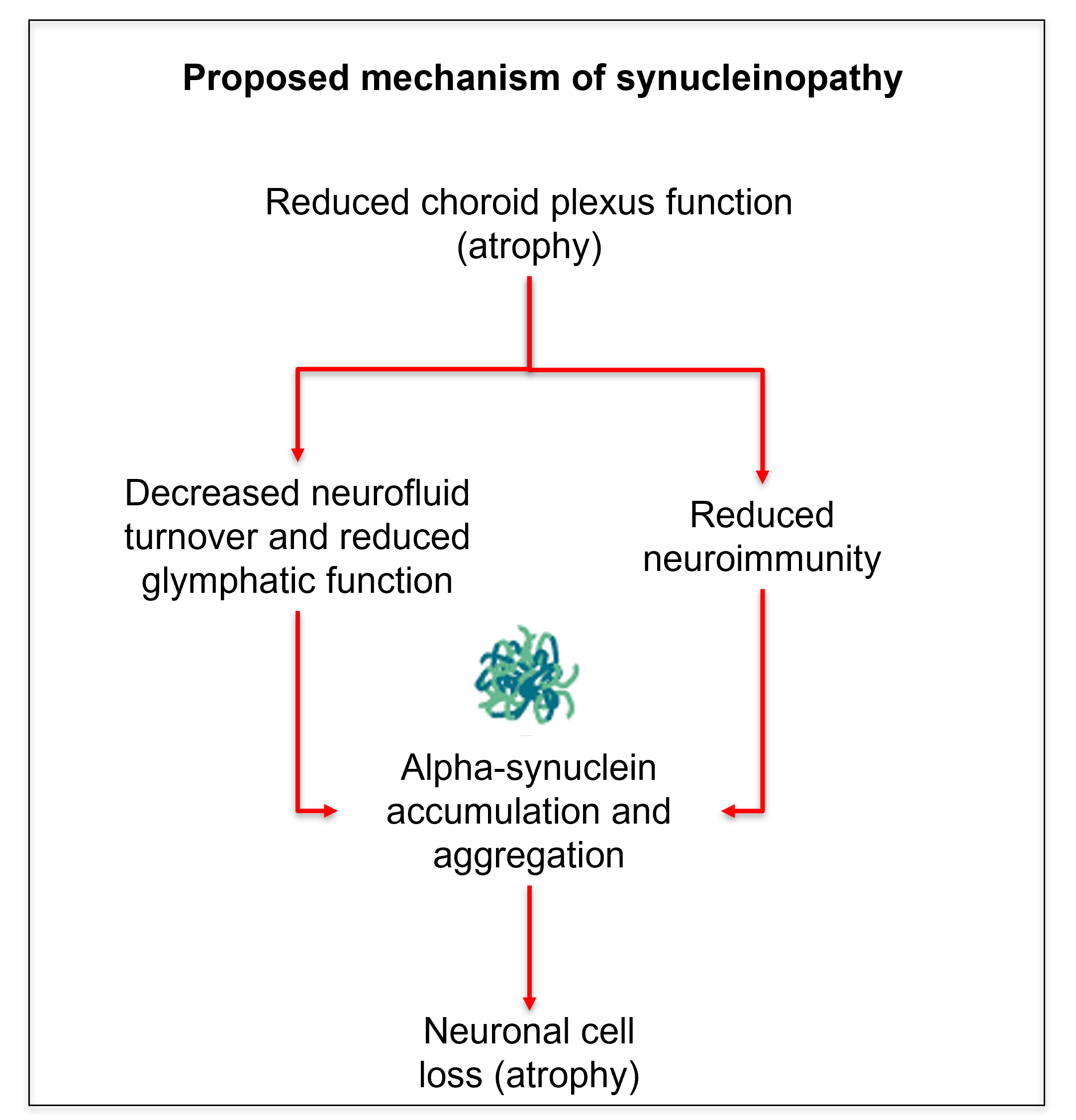
Accumulation and aggregation of misfolded α-synuclein protein is the hallmark of neurodegenerative α-synucleinopathies, including Parkinson's disease, dementia with Lewy bodies, multiple system atrophy, and, at the premanifest stage, isolated Rapid Eye Movement sleep behavior disorder (iRBD).1 Yet, it is still unclear which brain mechanisms lead to or exacerbate the brain accumulation of α-synuclein, especially at premanifest stages, when neuroprotective/neuromodulating treatment would be most effective. Accumulating evidence in the last decade2-10 indicates that the choroid plexus (ChP) is a critical player in neurosecretory/glymphatic and neuroimmune mechanisms, with its dysfunction often linked to protein aggregation and neuroinflammation. For instance, changes in ChP volume have been reported in aging, neuroinflammatory disease,10 and Alzheimer’s disease2-9. Based on these studies, here we hypothesized a similar mechanism for neurodegenerative α-synucleinopathies, in which structural changes of the ChP would be detectable at premanifest synucleinopathy stages, potentially affecting α-synuclein accumulation/aggregation in the brain (Figure 1).Purpose
Using high-spatial resolution 7 Tesla MRI, our aim was to: (i) test the hypothesis that the choroid plexus displays morphological changes in premanifest synucleinopathy, indicating decreased neurosecretory/glymphatic and neuroimmune function; (ii) create an original probabilistic atlas of the choroid plexus in the lateral ventricles.Materials and Methods
Cohorts: Twelve patients with premanifest synucleinopathy (iRBD) (age 68±5.70 years, 1f/11m) and twelve age/gender-matched controls (age 65.6±6.2 years, 1f/11m) underwent 7 Tesla MRI under IRB-approval. iRBD patients display REM sleep without atonia and dream-related behaviors11, and face up to a 90% risk of developing a neurodegenerative α-synucleinopathies within 14 years from diagnosis12; thus, they are considered a premanifest synucleinopathy.13 Data acquisition: T1-weighted multi-echo MPRAGE (MEMPRAGE): voxel-size:0.75mm-isotropic; repetition-time(TR):2.53s; echo-times(TEs):[2.39,5.62ms]ms; inversion-time:1.1s; flip-angle(FA):7°; field-of-viewFOV:240×240×240mm3; bandwidth:332Hz/pixel; GRAPPA-factor:3; slice-orientation:sagittal; slice-acquisition-order: anterior-posterior; acquisition-time: 6′34′′. T2*-weighted echo-planar-images (EPIs): Functional gradient-echo EPIs were acquired with the subjects at rest with their eyes closed. Voxel-size: 1.1mm-isotropic; TR:2.5s; TE:32ms; FA:75°; matrix size:180×240; GRAPPA-factor:3; nominal-echo-spacing:0.82ms; readout bandwidth:1488Hz; N.slices:123; slice orientation:sagittal; slice-acquisition-order:interleaved; acquisition-time9’25”. Field-map (acquired for distortion-correction): Voxel-size:2.0mm; matrix-size:116x132; readout-bandwidth:1515 Hz/pixel; N. slices:80; slice-orientation:sagittal; slice-acquisition-order:interleaved; TEs[:3.00 4.02]ms; TR:570.0ms; FA:36°; simultaneous-multi-slice-factor:3; phase-encoding-direction:anterior-posterior; acquisition-time:2’15’’. Data analysis: a) Preprocessing: The T1-weighted image was rotated to standard-orientation, cropped in z-axis to cover the brain until at least 2-3 cm below the obex, and bias-field corrected. T2*-weighted images were corrected for physiological noise (RETROICOR), slice-timing corrected, reoriented, distortion- (FSL, using the field-map) and motion-corrected (Freesurfer). The time-averaged image was bias-field corrected (SPM) and coregistered to the MEMPRAGE using affine and nonlinear (AFNI20) transformations. b) Evaluation of ChP volume and microstructure: The left and right ChP in the lateral ventricles were manually segmented on multi-contrast MRI, using both the T1-weighted image and the T2*-weighted image coregistered to the T1-weighted image. Volume (in mm3, normalized by the total intracranial-volume) and mean T2*-weighted and T1-weighted signal intensity of ChP were computed, and compared between cohorts using a two-sample t-test. c) Probabilistic atlas generation: Single-subject ChP segmentations were coregistered to stereotactic (Montreal Neurologica Institute, MNI) space using nonlinear (ANTs) registrations, and averaged across subjects to create a probabilistic atlas of the left and right ChP of the lateral ventricles.Results
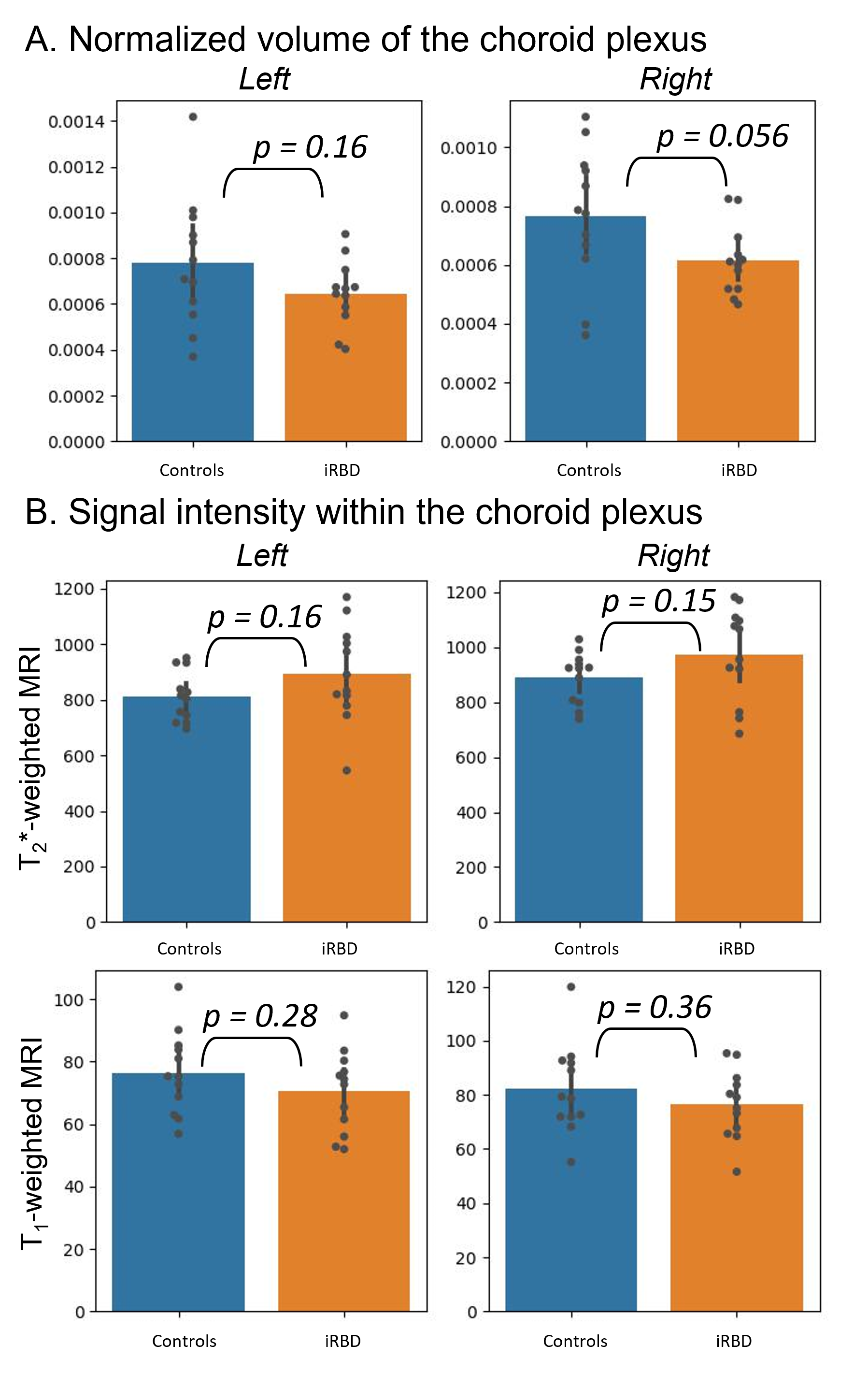
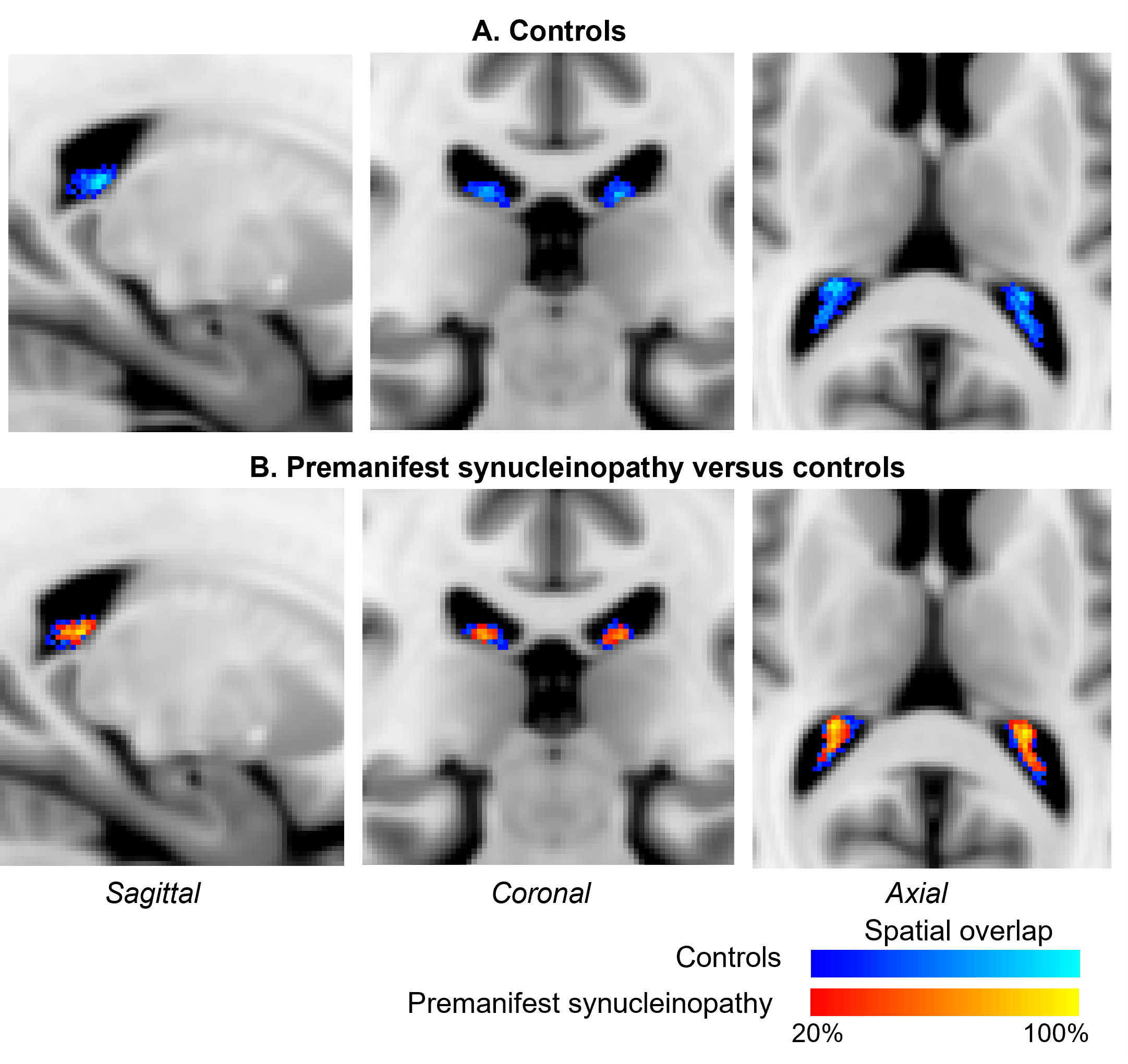
In premanifest synucleinopathy, we observed a significant decreased volume of the right ChP, with a similar trend for the left ChP. In addition, we found trends towards longer T2*-weighted mean values in both ChP, without significant changes in mean T1-weighted values (Figure 2). A probabilistic atlas of left and right ChP of the lateral ventricles was created in stereotactic space (Figure 3).Discussion and conclusion
In premanifest synucleinopathy, we found a decreased ChP volume, possibly related to ChP epithelial cell atrophy as in Alzheimer’s disease,9 which might underlie a decreased neuroimmune function and decreased neurofluid production in these patients. Crucially, a decreased neurofluid production directly relates to a decreased neurofluid turnover,2 increased neurofluid stasis, and thus reduced glymphatic clearance14 of brain waste and neurotoxic proteins. In addition, we found a trend towards increasing T2*-weighted values, which are compatible with atrophy and exclude the presence of major calcifications, which would have shortened the T2*-weighted values instead. These results shed light into future exploration of ChP as potential synucleinopathy biomarker. Further, we created an original preliminary probabilistic atlas of the choroid plexus of the lateral ventricles in living humans, in which ChP shrinking can be visually seen when comparing iRBD and controls. This atlas will be useful for the community as a prior to automatically identify the location of the choroid plexus in conventional and advanced MRI of living humans.Acknowledgements
NIH (NIA-R01AG063982); MJFF-022672 Award.References
- Stefanis L. α-Synuclein in Parkinson's disease. Cold Spring Harb Perspect Med. 2012;2(2):a009399.
- Serot J-M, Zmudka J, Jouanny P. A Possible Role for CSF Turnover and Choroid Plexus in the Pathogenesis of Late Onset Alzheimer’s Disease. JAD 2012;30:17–26.
- Badaut J, Ghersi-Egea J-F. The Choroid Plexus and Cerebrospinal Fluid System: Roles in Neurodegenerative Diseases. In: The Choroid Plexus and Cerebrospinal Fluid. Elsevier 2016; 129–54.
- González-Marrero I, Giménez-Llort L, Johanson CE, Carmona-Calero EM, Castañeyra-Ruiz L, Brito-Armas JM, et al. Choroid plexus dysfunction impairs beta-amyloid clearance in a triple transgenic mouse model of Alzheimer’s disease. Front Cell Neurosci 2015;9:17.
- Miklossy J, Kraftsik R, Pillevuit O, Lepori D, Genton C, Bosman FT. Curly fiber and tangle-like inclusions in the ependyma and choroid plexus--a pathogenetic relationship with the cortical Alzheimer-type changes? J Neuropathol Exp Neurol 1998;57:1202–12.
- Wen GY, Wisniewski HM, Kascsak RJ. Biondi ring tangles in the choroid plexus of Alzheimer’s disease and normal aging brains: a quantitative study. Brain Res 1999;832:40–6.
- Serot JM Bene MC Faure GC. Choroid plexus, aging of the brain, and Alzheimer's disease. Front Biosci 2003;8:s515–21.
- Serot JM Foliguet B Bene MC , et al. . Choroid plexus and ageing in rats: A morphometric and ultrastructural study . Eur J Neurosci 2001;14:794–8.
- Serot J. M., Bene M. C., Foliguet B., Faure G. C. (2000). Morphological alterations of the choroid plexus in late-onset Alzheimer’s disease. Acta Neuropathol. 99;105–108.
- Martirosian V, Julian A, Neman J. The Role of the Choroid Plexus in the Pathogenesis of Multiple Sclerosis. In: The Choroid Plexus and Cerebrospinal Fluid. Elsevier 2016;103–27.
- Schenck CH, Bundlie SR, Ettinger MG, et al. Chronic behavioral disorders of human REM sleep: a new category of para- somnia. Sleep 1986;9(2):293-308.
- Galbiati A, Verga L, Giora E, Zucconi M, Ferini-Strambi L. The risk of neurodegeneration in REM sleep behavior disorder: A systematic review and meta-analysis of longitudinal studies. Sleep Med Rev. 2019;43:37-46.
- Iranzo A, Fernández-Arcos A, Tolosa E, Serradell M, Molinuevo JL, Valldeoriola F, et al. Neurodegenerative disorder risk in idiopathic REM sleep behavior disorder: study in 174 patients. PLoS One 2014;9:89741.
- Taoka T, Naganawa S. Neurofluid Dynamics and the Glymphatic System: A Neuroimaging Perspective. Korean J Radiol. 2020;21(11):1199-1209
Figures