3036
Neural correlates of anxiety in rhesus macaques exposed to early social adversity: A longitudinal DTI study across adolescence.1Institut des sciences cognitives Marc Jeannerod (ISC) , CNRS, LYON, France, 2Claude Bernard University, Lyon 1, LYON, France, 3CERMEP - Centre d'Etude et de Recherche Multimodal Et Pluridisciplinaire en imagerie du vivant, LYON, France
Synopsis
Keywords: White Matter, Diffusion Tensor Imaging
Using diffusion tensor imaging (DTI) and behavioral observation, this study investigated the longitudinal effects of early social adversity (ESA) on risk for anxiety and the neural mechanisms underlying this relationship across adolescence.
Diffusion MRI data were collected from 21 macaques comprising two groups that differed in exposure to ESA; mother-reared and nursery-reared. Each monkey was scanned at 2.5, 3.5 and 4.5 years old, with anxiety-like behavior assessed at each time-point.
Results suggest that ESA has long-term effects on WM fractional anisotropy (FA) in several regions, with differences in frontolimbic and frontostriatal WM tracts modulating the relationship between ESA and anxiety .
Introduction
Early social adversity (ESA) has been linked to poor outcomes in numerous developmental domains, including risk for anxiety disorders [1]. However, currently we know little about the neural mechanisms that may underlie the ESA-anxiety relationship across time. Accordingly, the aim of this study was to examine the longitudinal relationship between anxiety symptoms and the white matter development of brain circuitry related to anxiety disorders and known to be impacted by ESA. Specifically, we wanted to investigate brain development across adolescence, which is the key period of risk for pathological anxiety. To achieve this, we assessed two groups of macaques (one nursery-reared/ESA-exposed; one mother-reared/non-ESA exposed) at three time-points corresponding approximately to pre-adolescence (2.5 years), adolescence (3.5 years), and early adulthood (5 years). At each time-point, diffusion MRI (dMRI) was obtained to examine white matter microstructure, and videos of animals were recorded to identify anxiety-like behavior. We hypothesized that WM integrity in frontolimbic and frontostriatal tracts (i.e. circuitry involved in emotion regulation, motivation, and reward) would modulate the relationship between ESA and anxiety.Materials and methods
The sample (n = 21) was composed of two groups of macaques; 11 raised by their mothers (mother-reared) and 10 raised in a nursery of peers by human caregivers (nursery-reared). In each of the two groups, approximately half of the monkeys were male and the other half were female.A 3D multi-shot Echo-planar (3D msEPI) pulse sequence [2] with an isotropic spatial resolution of 0.8 mm3 was used to collect diffusion-weighted (DW) brain images with b =1000 s/mm² applied across 46 non-collinear diffusion directions and 2 non-DW images ( b=0 s/mm²) for each macaque at each time-point at 3T.
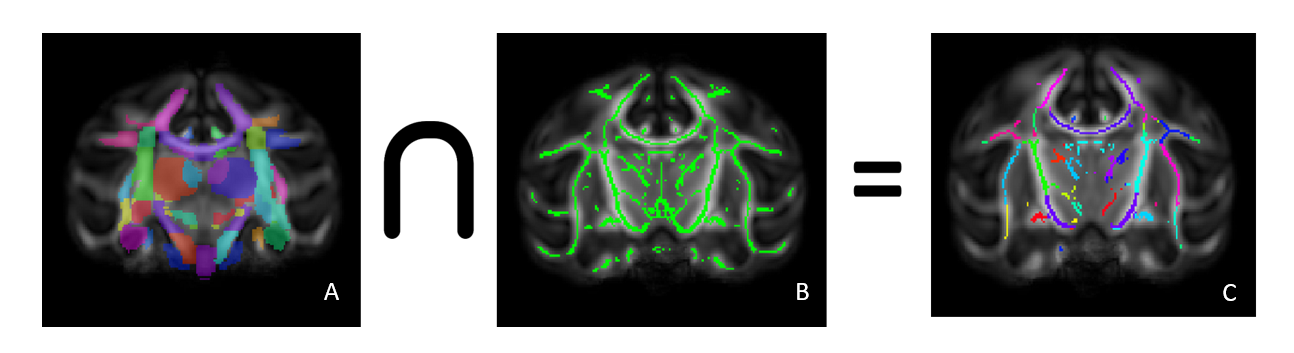
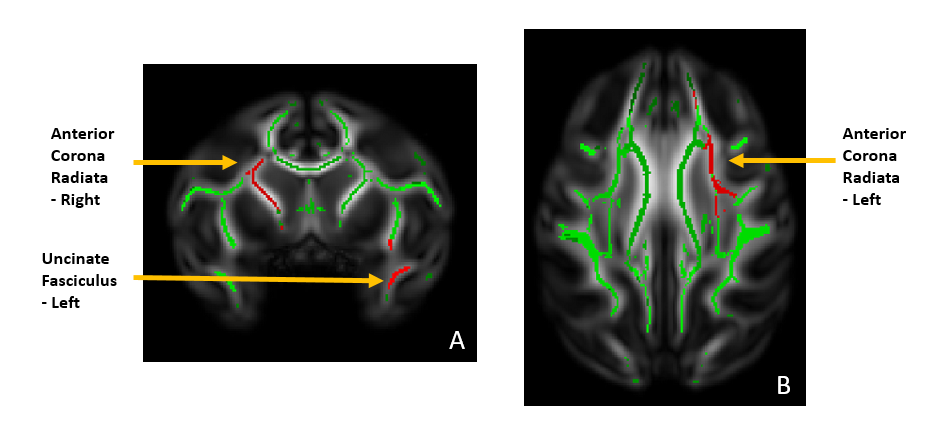
dMRI preprocessing included denoising, distortions / eddy currents correction and DTI-maps (fractional anisotropy (FA), radial diffusivity (RD), axial diffusivity (AD), mean diffusivity (MD)) computing. A study specific template was created using all FA images across time-points and subjects with ANTS optimal template construction [3].WM skeletons of all the macaques were defined using tract-based spatial statistics (TBSS); the first elastic registration step was replaced by a tensor registration method [4] that incorporates all tensor information and improves alignment with the model using DTI-TK [5]. Overlaying the ONPRC18 multimodal macaque MRI atlas [6] on the TBSS skeleton images allowed the analysis of skeleton regions of interest by calculating the mean DTI metric of these regions and running statistics on them. Based on previous literature [7], we selected five ROIs for analysis; uncinate fasciculus (left/right), fornix, and anterior corona radiata (left/right).
Behavioral observations reflecting anxiety in macaques : self-scratching, self-grooming, yawns, and body shakes were coded using the focal sampling method [8] . For each year, the frequency of these behaviors were averaged across recording sessions to give the final anxiety score.
A mixed modeling approach was used for all analyses. For exploratory analyses of DTI metrics on the skeleton ROIs we used group and time-point as fixed effects, and subject-specific intercepts as a random effect. Negative binomial generalized mixed models were used to examine how ESA was linked to anxiety; group, time point and FA were included as fixed effects, and subject-specific intercepts as a random effect. Sex was controlled for in all models.
Results
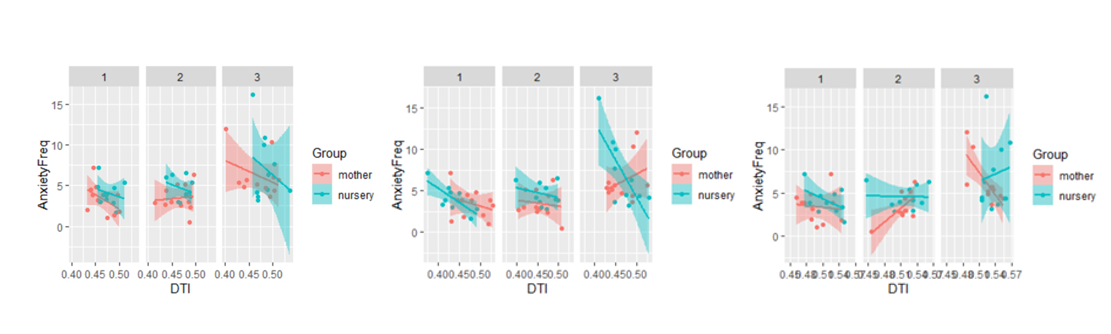
Exploratory statistics revealed that in the fornix, FA was significantly higher in the mother-reared group in the first year, but no significant difference between the groups was revealed in the second and third years. RD was significantly higher for the mother-reared group in the left uncinate fasciculus although increasing over the years for both groups whereas for the anterior corona radiata it’s decreasing over the years but still significantly higher for the mother-reared group.Anxiety was significantly greater in the nursery-reared versus mother-reared groups across time, with anxiety also increasing across groups with time. FA in all tracts of interest significantly modulated the relationship between ESA and anxiety. For the right uncinate fasciculus and fornix, a group by FA by time-point interaction was revealed; in the mother-reared group, a negative relationship between FA (i.e. more FA linked to less anxiety) decreased over time, whereas this negative relationship persisted in the nursery-reared group. A significant group by FA by time-point interaction was also revealed for the right anterior corona radiata; at time-points one and three, there was a negative relationship between FA and anxiety in the mother-reared group, but a positive relationship at time-point two. In the nursery-reared group there was a negative relationship at time-point one, but a positive relationship at time-point three.
Discussion
Results from this study revealed widespread effects of ESA on development of WM microstructure across adolescence. As predicted, longitudinal relationships across this period between ESA and anxiety were found, with this relationship modulated by both frontolimbic and frontostriatal tracts. These findings suggest that while greater WM integrity in certain circuitry may be protective against anxiety in the context of ESA, this may also be linked to increased risk.Conclusion
These findings provide novel insights in the long-term trajectories of anxiety after ESA, and the neural mechanisms that underlie this relationship. Such knowledge will be essential for identifying individuals most at risk and when, and thus for the development of more effective treatments and early interventions after exposure to ESAAcknowledgements
No acknowledgement found.References
1. Wade M, Parsons J, Humphreys KL, McLaughlin KA, Sheridan MA, Zeanah CH, Nelson CA, Fox NA. The Bucharest Early Intervention Project: Adolescent mental health and adaptation following early deprivation. Child Development Perspectives. 2022 Sep;16(3):157-64.
2. S. Tounekti, T. Troalen, Y. Bihan-Poudec, M. Froesel, F. Lamberton, V. Ozenne, J. Cléry, N. Richard, M. Descoteaux, S. Ben Hamed, B. Hiba,High-resolution 3D diffusion tensor MRI of anesthetized rhesus macaque brain at 3T,NeuroImage, Volume 181, 2018, Pages 149-161, ISSN 1053-8119.
3. Brian B. Avants, Nick Tustison and Hans Johnson3 Advanced Normalization Tools (ANTS) , Release 2.x, page 21, July 10, 2014 University of Pennsylvania.
4.Keihaninejad S, Zhang H, Ryan NS, Malone IB, Modat M, Cardoso MJ, Cash DM, Fox NC, Ourselin S. An unbiased longitudinal analysis framework for tracking white matter changes using diffusion tensor imaging with application to Alzheimer's disease. Neuroimage. 2013 May 15;72:153-63. doi: 10.1016/j.neuroimage.2013.01.044. Epub 2013 Jan 28. PMID: 23370057.
5. Zhang, H., Avants, B.B, Yushkevich, P.A., Woo, J.H., Wang, S., McCluskey, L.H., Elman, L.B., Melhem, E.R., Gee, J.C., High-dimensional spatial normalization of diffusion tensor images improves the detection of white matter differences in amyotrophic lateral sclerosis, IEEE Transactions on Medical Imaging, 26(11):1585-1597, November 2007.
6. Alison R. Weiss, Zheng Liu, Xiaojie Wang, William A. Liguore, Christopher D. Kroenke, Jodi L. McBride, The macaque brain ONPRC18 template with combined gray and white matter labelmap for multimodal neuroimaging studies of Nonhuman Primates,NeuroImage,Volume 225,2021,117517,ISSN 1053-8119.
7. eeBick J, Zhu T, Stamoulis C, Fox NA, Zeanah C, Nelson CA. Effect of early institutionalization and foster care on long-term white matter development: a randomized clinical trial. JAMA pediatrics. 2015 Mar 1;169(3):211-9.
8. Coleman K, Pierre PJ. Assessing anxiety in nonhuman primates. ILAR journal. 2014 Jan 1;55(2):333-46.
Figures