1457
Blockage of CSF Outflow via Nasal Lymphatic Pathway in Rats after Deep Cervical Lymph Node Ligation Observed Using Intrathecal Gd-based MR Imaging1Department of Diagnostic Imaging, Faculty of Medicine and Graduate School of Medicine, Hokkaido University, Sapporo, Japan, 2Department of Dental Radiology, Hokkaido University Hospital, Sapporo, Japan, 3Department of Radiology, Faculty of Dental Medicine, Hokkaido University, Sapporo, Japan, 4Department of Diagnostic and Interventional Radiology, Hokkaido University Hospital, Sapporo, Japan, 5Global Center for Biomedical Science and Engineering, Faculty of Medicine, Hokkaido University, Sapporo, Japan
Synopsis
Keywords: Neurofluids, Neurofluids, Glymphatic System
Previous reports showed that deep cervical lymph nodes (DCLNs) receive cerebrospinal fluid (CSF) outflow. DCLN ligation resulted in intracranial accumulation of waste proteins due to impaired CSF outflow; however, changes in extracranial outflow after DCLN ligation have not been directly observed. We examined extracranial CSF outflow in rats after DCLN ligation by using intrathecal gadolinium (Gd)-based dynamic contrast-enhanced MRI. DCLN ligation blocked CSF- tracer outflow into the nasal cavity. A weak trend towards CSF-tracer retention in the ventral cistern was also observed. These results suggest that DCLN ligation affects CSF outflow in rodents.Introduction
The transnasal pathway is a major cerebrospinal fluid (CSF) outflow route in rodents.1-3 Recent studies have also reported the existence of an efflux pathway via meningeal lymphatic vessels.4 Connections between deep cervical lymph nodes (DCLNs) and these efflux pathways may serve as relay points for CSF and solute outflow.2–8 Surgical ligation of DCLNs accelerates the intracranial accumulation of amyloid-β or α-synuclein by blocking the outflow of CSF, and exacerbates the symptoms of Alzheimer’s and Parkinson’s diseases in mouse models.7,8 Although such pathological effects have been reported, changes in extracranial CSF outflow after ligation of DCLNs have not been directly observed. In this study, we examined whether DCLN ligation changes the extracranial outflow of CSF tracers using a rat model of Gd-based dynamic contrast-enhanced (DCE) MRI.Material and Methods
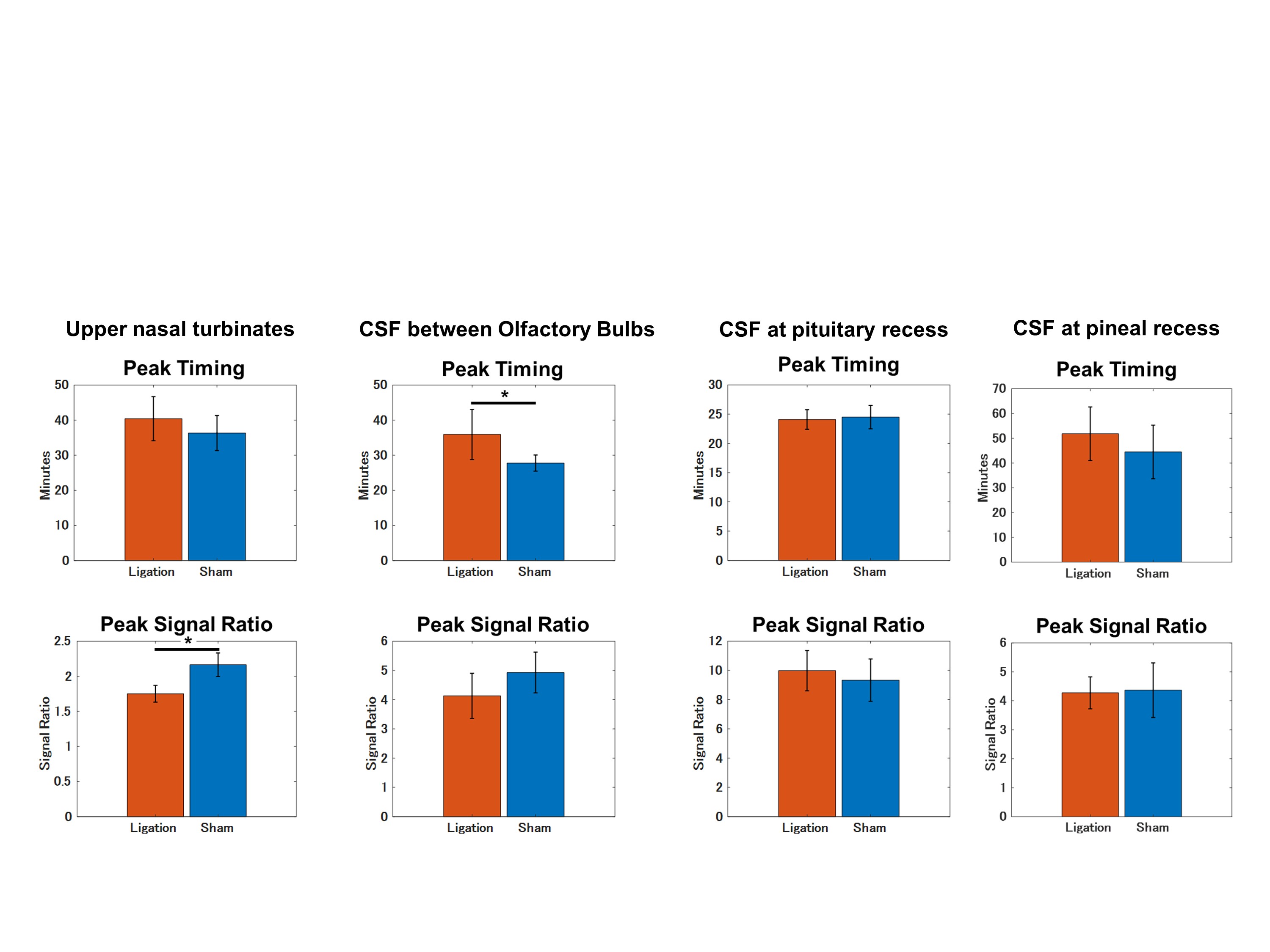
The Animal Study Committee of our institute approved all study experiments. Male Wistar 7-10-week-old rats were used in the study. Twelve rats were divided into two groups: a bilateral DCLN-ligation group (n = 6) and a sham operation group (n = 6). After placing an intrathecal injection tube in the cisterna magna, DCE MRI was conducted using a 3.0-T clinical scanner (Magnetom Prisma, Siemens, Germany) with a custom-made 8-channel phased array receiver coil (Takashima Seisakusho, Japan). Scans were acquired using 3D T1 weighted volumetric interpolated breath-hold examination sequences (VIBE) with the following parameters: TR, 8.72 ms; TR, 3.6 ms; flip angle, 14 deg; acquisition resolution, 0.4 × 0.4 × 0.2 mm; reconstruction resolution, 0.2 × 0.2 × 0.2 mm; number of excitations, 1; scan time per phase, 147 s; number of repetitions, 43; and total scan time, 105 min 21 s. After acquiring three phases of pre-contrast images, the rats received an intrathecal injection of 10 mmol/L of gadolinium (Gd)-BTDO3A (Gadovist, Bayer HealthCare Japan, Japan) diluted in normal saline at 1.7 μL/min for 20 min using an MR-compatible microinjector, resulting in a total amount of 34 μL. After initiating intrathecal injection, 40 phases of 3D-T1 VIBE images were acquired. Details of the procedures are shown in Figure 1. Rectangle ROIs were manually placed on CSF space at the pituitary recess, between olfactory bulbs and the pineal recess. Polygonal ROIs were placed on upper nasal turbinates and soft tissue outside the bilateral jugular foramen. For each ROI, the signal ratio of each phase from the average of 3 pre-contrast phases was calculated, and average time-intensity curves (TICs) were generated for both groups and compared using 2-way repeated measures ANOVA. The peak signal ratios and peak timing of signal ratios were also compared using U-tests. A p-value of < 0.05 was considered significant.Results
Gd-BTDO3A injected from the cisterna magna spread through the CSF space rostrally over time and flowed out through the cribriform plate into the nasal cavity (Figure 2 and 3). The DCLN-ligation group demonstrated a lower signal ratio with statistical significance in the upper nasal turbinates in the early phases from 17.2 to 31.9 minutes after injection initiation (Figure 4). In the ligation group, we observed a trend towards a higher signal ratio in the CSF space at the pituitary recess in the delayed phase, but this was not statistically significant. No apparent signal change was observed in the soft tissue around the jugular foramen. The peak signal-ratio timing in the CSF was delayed at the level of the olfactory bulb in the DCLN-ligation group (p = 0.02), and we also observed a higher peak signal ratio in the upper nasal turbinates in the DCLN-ligation group (p = 0.002) (Figure 5).Discussion
Our results suggest that ligation of DCLNs blocks the nasal lymphatic pathway in rodents. This observation is consistent with previous studies proposing that intrathecally administered tracers mainly drain into the nasal cavity and travel through the peripheral lymphatic vessels of the nasopharynx to the DCLNs.1–3 Previous reports showed the contribution of meningeal lymphatics to CSF clearance, and several studies demonstrated an accumulation of Gd tracers in the lymphatic vessels around the jugular foramen using sequences sensitive to small amounts of Gd.3,4 In the present study, we did not detect this accumulation using the T1-VIBE sequence, in contrast to the clear observation of tracer outflow into the nasal cavity. This result indicated that in rats, the amount of CSF tracer flowing out from the jugular region was significantly smaller than that from the nasal pathway. In this study, a weak trend towards tracer retention in the basal CSF space during the late phases was observed. This might be explained by the blocked CSF outflow of the nasal lymphatic pathway. However, other compensatory mechanisms might exist, such as distribution into the brain or spinal subarachnoid space, because the observed difference was small. We were interested to elucidate the possible compensatory mechanisms; however, the tracer was not sufficiently distributed into the brain parenchyma under our injection conditions containing low Gd concentrations, (data not shown), and the spinal cord was outside the field of view.Conclusion
DCLN ligation blocks the nasal lymphatic pathway in rats. DCLN ligation may also cause solute retention in the basal CSF space.Acknowledgements
No acknowledgement found.References
1. Nagra G, Koh L, Zakharov A, et al. Quantification of cerebrospinal fluid transport across the cribriform plate into lymphatics in rats. Am J Physiol Regul Integr Comp Physiol. 2006;291(5):R1383-9.
2. Decker Y, Krämer J, Xin L, et al. Magnetic resonance imaging of cerebrospinal fluid outflow after low-rate lateral ventricle infusion in mice. JCI Insight. 2022;7(3):e150881.
3. Norwood JN, Zhang Q, Card D, et al. Anatomical basis and physiological role of cerebrospinal fluid transport through the murine cribriform plate. Elife. 2019;8:e44278.
4. Ahn JH, Cho H, Kim JH, et al. Meningeal lymphatic vessels at the skull base drain cerebrospinal fluid. Nature. 2019 Aug;572(7767):62-66.
5. Xue Y, Liu X, Koundal S, et al. In vivo T1 mapping for quantifying glymphatic system transport and cervical lymph node drainage. Sci Rep. 2020;10(1):14592.
6. Ma Q, Ineichen BV, Detmar M, et al. Outflow of cerebrospinal fluid is predominantly through lymphatic vessels and is reduced in aged mice. Nat Commun. 2017;8(1):1434.
7. Zou W, Pu T, Feng W, Lu M, et al. Blocking meningeal lymphatic drainage aggravates Parkinson's disease-like pathology in mice overexpressing mutated α-synuclein. Transl Neurodegener. 2019;8(1):7.
8. Wang L, Zhang Y, Zhao Y, et al. Deep cervical lymph node ligation aggravates AD-like pathology of APP/PS1 mice. Brain Pathol. 2019;29(2):176-192.
Figures
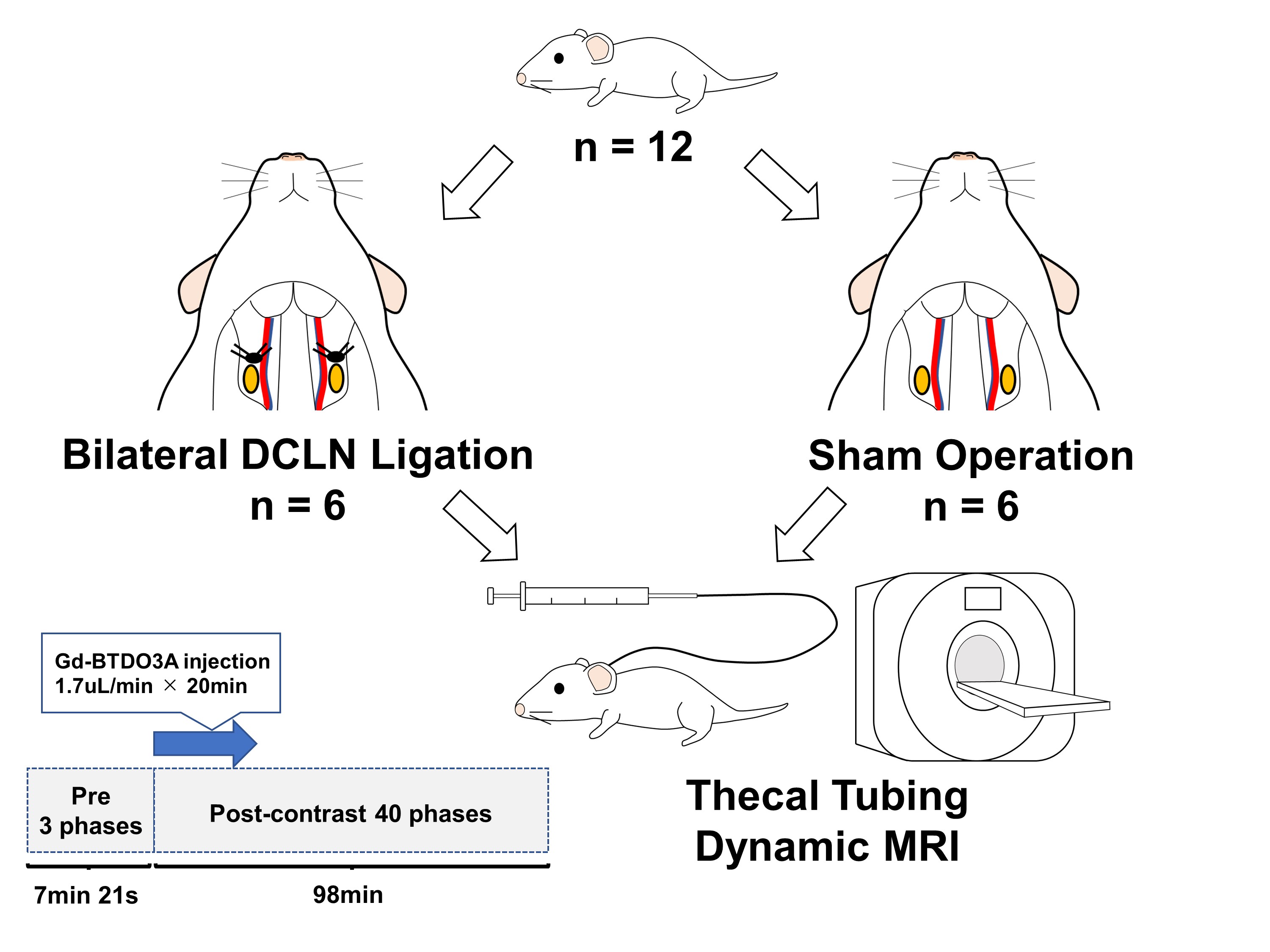
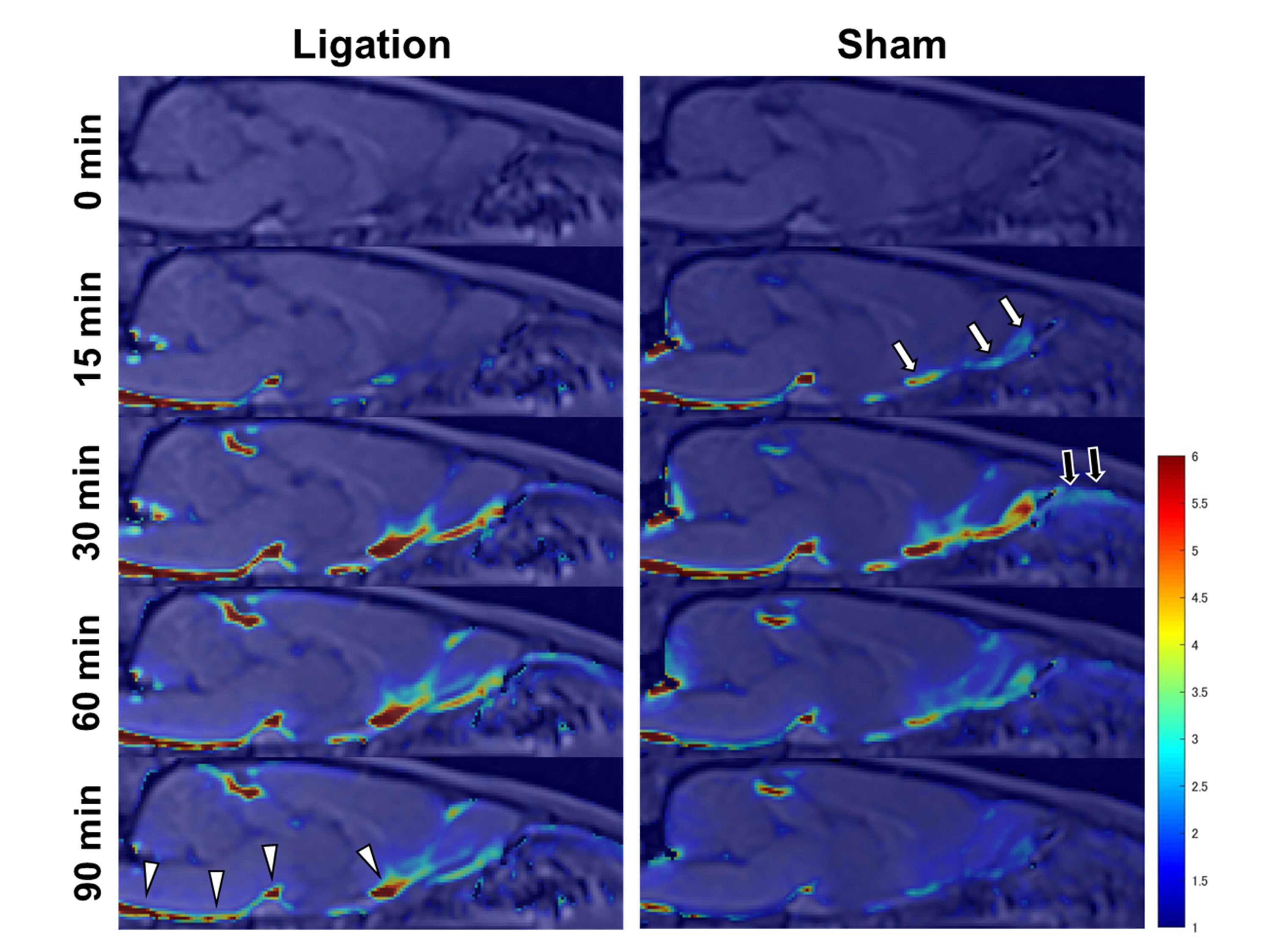
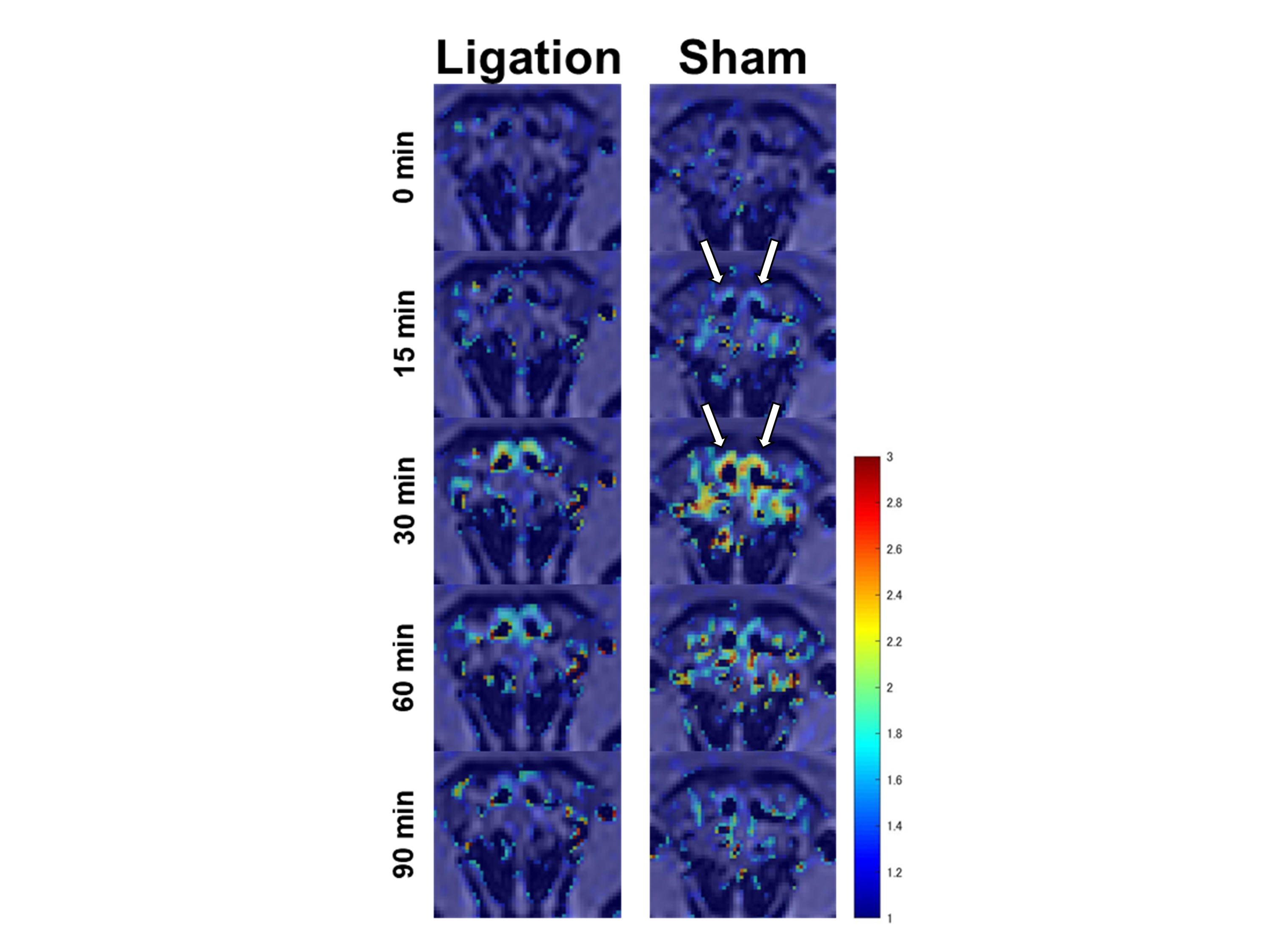
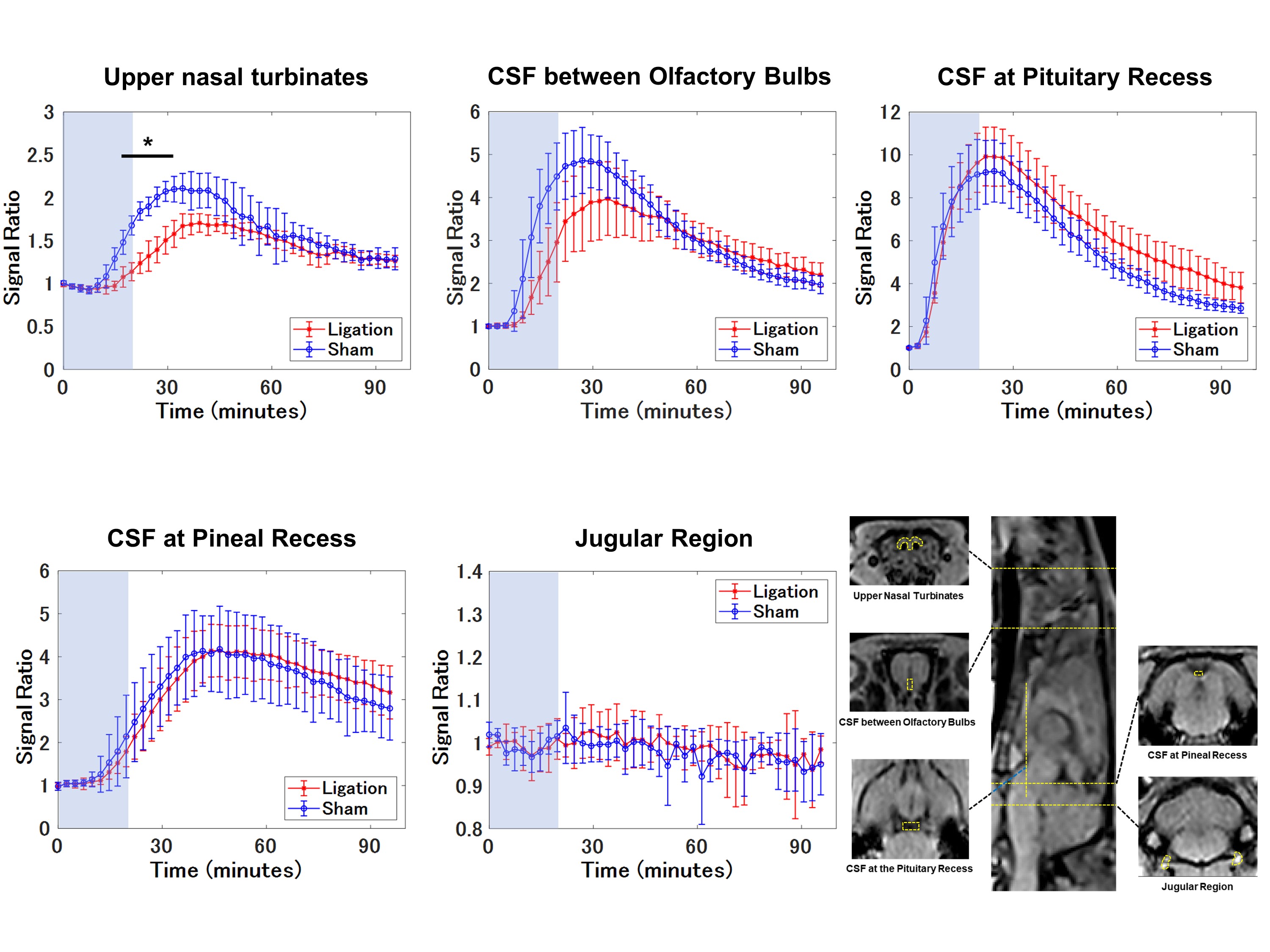
Figure 4. Average time intensity curves (TIC’s).
The average TIC’s and methods for setting each of five ROI’s are shown. The DCLN-ligation group shows a lower signal ratio in the upper nasal turbinates in the early phases. In the CSF at the pituitary recess, a weak trend toward a higher signal ratio is observed in the delayed phases. Data are presented as the mean ± standard deviation. The light blue area represents an intrathecal injection duration of 20 min. * Indicates a statistically significant difference between the two groups (p < 0.05, two-way repeated-measures ANOVA).