1357
In-vivo imaging of the human thalamus: a comprehensive evaluation of structural imaging approaches for thalamic nuclei differentiation at 7T
Cristina Sainz Martinez1,2, José P. Marques3, Gabriele Bonanno4,5,6, Tom Hilbert4,7,8, Constantin Tuleasca8,9,10, Meritxell Bach Cuadra2,7, and João Jorge1
1CSEM - Swiss Center for Electronics and Microtechnology, Bern, Switzerland, 2CIBM Center for Biomedical Imaging, Lausanne, Switzerland, 3Donders Centre for Cognitive Neuroimaging, Radboud University, Nijmegen, Netherlands, 4Advanced Clinical Imaging Technology, Siemens Healthineers International AG, Bern, Switzerland, 5Translational Imaging Center (TIC), Swiss Institute for Translational and Entrepreneurial Medicine (SITEM), Bern, Switzerland, 6Magnetic Resonance Methodology, Institute of Diagnostic and Interventional Neuroradiology, University of Bern, Bern, Switzerland, 7Department of Radiology, Lausanne University Hospital (CHUV) and University of Lausanne (UNIL), Lausanne, Switzerland, 8Signal Processing Laboratory 5 (LTS-5), École Polytechnique Fédérale de Lausanne, Lausanne, Switzerland, 9Department of Clinical Neurosciences, Neurosurgery Service and Gamma Knife Center, Centre Hospitalier Universitaire Vaudois (CHUV), Lausanne, Switzerland, 10Faculty of Biology and Medicine, University of Lausanne (UNIL), Lausanne, Switzerland
1CSEM - Swiss Center for Electronics and Microtechnology, Bern, Switzerland, 2CIBM Center for Biomedical Imaging, Lausanne, Switzerland, 3Donders Centre for Cognitive Neuroimaging, Radboud University, Nijmegen, Netherlands, 4Advanced Clinical Imaging Technology, Siemens Healthineers International AG, Bern, Switzerland, 5Translational Imaging Center (TIC), Swiss Institute for Translational and Entrepreneurial Medicine (SITEM), Bern, Switzerland, 6Magnetic Resonance Methodology, Institute of Diagnostic and Interventional Neuroradiology, University of Bern, Bern, Switzerland, 7Department of Radiology, Lausanne University Hospital (CHUV) and University of Lausanne (UNIL), Lausanne, Switzerland, 8Signal Processing Laboratory 5 (LTS-5), École Polytechnique Fédérale de Lausanne, Lausanne, Switzerland, 9Department of Clinical Neurosciences, Neurosurgery Service and Gamma Knife Center, Centre Hospitalier Universitaire Vaudois (CHUV), Lausanne, Switzerland, 10Faculty of Biology and Medicine, University of Lausanne (UNIL), Lausanne, Switzerland
Synopsis
Keywords: Multi-Contrast, High-Field MRI, Thalamus, Nuclei, 7 Tesla, QSM
The ability to non-invasively image the thalamus and its different nuclei would be highly valuable to neuroscience and neuroradiology, but has remained challenging. Here, we initiated a comprehensive practical review of recent thalamic imaging approaches at 7 Tesla, based on T1, T2, T2* and susceptibility properties. These were all acquired on the same in-vivo brain, to avoid anatomical variability confounds. The images were qualitatively compared to histological atlases. Upon systematic assessment, QSM and GM/WM-optimized MP2RAGE proved the most valuable to differentiate specific nuclei. To our knowledge, this is the most comprehensive evaluation to date of thalamic imaging modalities at 7T.Introduction
The thalamus is a subcortical brain structure of major functional importance, which is anatomically subdivided in smaller nuclei with specialized functions. Alterations in function and/or structure of certain nuclei have been linked to disorders such as schizophrenia1 and multiple sclerosis2, and thus could provide valuable future imaging biomarkers; several nuclei have already been identified as effective clinical intervention targets in disorders such as drug-resistant tremor3,4 and epilepsy5,6. Unfortunately, the ability to non-invasively visualize the different nuclei in-vivo has remained considerably limited in the past, with conventional T1 and T2-weighted contrasts yielding little to no distinguishable features, and diffusion modalities only discriminating between larger nuclei groups7.However, recent efforts at ultra-high field, using mainly magnetic susceptibility8,9 and T1-weighted contrasts10,11, have shown promising unprecedented capabilities for thalamic nuclei mapping, in some cases able to distinguish even smaller nuclei such as the ventral-intermediate (Vim) nucleus12. Considering these recent findings, it is now highly pertinent to systematically evaluate the available options and determine the most effective solutions to image specific nuclei.
In this work, we have initiated a comprehensive practical review of recent thalamic imaging approaches at 7 Tesla. We covered diverse modalities based on T1, T2 and T2* properties (including quantitative susceptibility mapping (QSM)), with both conventional and “contrast-focusing” approaches. Crucially, these were acquired from the same individual brain, to allow direct comparisons across modalities without anatomical variability confounds. To provide anatomical references, the images were qualitatively compared to well-established thalamic atlases.
Methods
Data acquisition: The acquisitions were performed at 7T (MAGNETOM Terra, Siemens Healthcare, Erlangen, Germany) with a single-channel transmit/32-channel receive head coil (Nova Medical, Wilmington MA, USA), from a healthy 26-yo adult. The following research application sequences were acquired, split over several sessions (total TA≈2.3h):- Conventional MP2RAGE13 (TI1/TI2=800/2700ms), which yielded a uniform T1-weighted image (T1w) and T1 map (T1map);
- MPRAGE with TI/TR=680/6000ms, for a white matter-nulled image (WMnull)11
- MP2RAGE optimized for gray-to-white matter contrast (GWMopt)10 (TI1/TI2=700/1600ms);
- Five-echo 3D-GRE (ΔTE=5.2ms), repeated for 9 head orientations, which provided R2* (R2*map) and QSM (estimated using COSMOS14);
- 3D-GRE optimized for Susceptibility-Weighted Imaging (SWI) (TE=20ms)12;
- 3D-variable flip-angle (VFA) TSE with TF=96, TE/TR=146/2650ms, for a conventional T2-weighted image (T2w);
- Inversion-recovery 3D-VFA TSE (IRTSE)15 (TF=80, TI/TE/TR=500/148/5330ms).
Processing: All images were registered to WMnull (Fig.1a). To investigate the correspondence between image features and expected thalamic anatomy, two histological atlases were aligned with the data:
- THOMAS16, based on a simplified version of the Morel atlas17, fit to WMnull (Fig.1b);
- Schaltenbrand18, specifically slice z=3.5mm, which contains the Vim and other nuclei (Fig.1c).
Results
Morel-based atlas: The data confirmed that conventional contrasts, namely T1w and T2w, provide limited information in the thalamus (Fig.2). In contrast, susceptibility-based modalities, especially QSM, were remarkably informative – e.g., enabling the distinction between the Ventral Posterior Lateral (VPL), Ventral Lateral Posterior-Inferior (VLP-Inf) and Ventral Lateral anterior (VLa) nuclei (Fig.2-Slice ii, Fig.3). The optimized T1-weighted sequences, especially GWMopt, offered the clearest distinctions across the medial-lateral direction – e.g., between the Mediodorsal-Parafascicular (MD-Pf) and Ventral Lateral Posterior-superior (VLP-sup) nuclei. Importantly, in more superior regions (Fig.2-Slice iii), the susceptibility-based modalities, especially SWI, were sensitive to large veins, which hindered nuclei visualization. Some superior-anterior nuclei such as the Ventral Lateral Posterior-superior (VLP-sup), Anterior Ventral (AV) and Ventral Anterior (VA) remained challenging to differentiate with all tested modalities (Fig.2-Slice iii).Schaltenbrand atlas: The observed anatomical features were generally in good agreement with those of Morel (Fig.4). SWI and especially QSM offered the clearest and most complete differentiations, including e.g., the Vim. Nonetheless, GWMopt offered the clearest contrast between the Parafascicular (Pf) and Centromedian (Cm) nuclei, for example.
Modality comparison: Together, QSM and GWMopt provided the clearest distinctions between almost every nucleus pair considered (Fig.3,4,5). Other T1-based modalities (T1w,T1map,WMnull) appeared sensitive to roughly the same features as GWMopt, but with poorer SNR and/or contrast. SWI showed the same trend with respect to QSM, but was more affected by perturbations such as from veins; R2*map lacked contrast. The SNR of T2w and IRTSE was too low to allow conclusive observations, and could potentially benefit from further optimization.
Conclusion
To our knowledge, this study presents the most comprehensive qualitative evaluation to date of existing modalities for thalamic imaging at 7T, all applied to the same in-vivo brain. The data indicate that QSM and GM/WM-optimized MP2RAGE are the most valuable tools currently available, allowing clear delineations of numerous thalamic nuclei. These findings will be consolidated in future work with additional subjects, and potentially more quantitative evaluation metrics.Acknowledgements
This work was funded by the Swiss National Science Foundation (SNSF) through grant PZ00P2_18590, and supported by the Swiss Center for Electronics and Microtechnology (CSEM) and the CIBM Center for Biomedical Imaging, a Swiss research center of excellence founded and supported by CHUV, UNIL, EPFL, UNIGE, HUG and the Leenaards and Jeantet Foundations.References
- N. D. Woodward, H. Karbasforoushan, and S. Heckers, ‘Thalamocortical Dysconnectivity in Schizophrenia’, AJP, vol. 169, no. 10, pp. 1092–1099, Oct. 2012, doi: 10.1176/appi.ajp.2012.12010056.
- N. Bergsland et al., ‘Thalamic Nuclei Volumes and Their Relationships to Neuroperformance in Multiple Sclerosis: A Cross‐Sectional Structural MRI Study’, Magnetic Resonance Imaging, vol. 53, no. 3, pp. 731–739, Mar. 2021, doi: 10.1002/jmri.27389.
- A. L. Benabid et al., ‘Chronic electrical stimulation of the ventralis intermedius nucleus of the thalamus as a treatment of movement disorders’, Journal of Neurosurgery, vol. 84, no. 2, pp. 203–214, Feb. 1996, doi: 10.3171/jns.1996.84.2.0203.
- A. Kooshkabadi, L. D. Lunsford, D. Tonetti, J. C. Flickinger, and D. Kondziolka, ‘Gamma Knife thalamotomy for tremor in the magnetic resonance imaging era: Clinical article’, JNS, vol. 118, no. 4, pp. 713–718, Apr. 2013, doi: 10.3171/2013.1.JNS121111.
- A. Sitnikov, Y. Grigoryan, and L. Mishnyakova, ‘Bilateral stereotactic lesions and chronic stimulation of the anterior thalamic nuclei for treatment of pharmacoresistant epilepsy’, Surg Neurol Int, vol. 9, no. 1, p. 137, 2018, doi: 10.4103/sni.sni_25_18.
- J. F. Kerrigan et al., ‘Electrical Stimulation of the Anterior Nucleus of the Thalamus for the Treatment of Intractable Epilepsy’, Epilepsia, vol. 45, no. 4, pp. 346–354, Apr. 2004, doi: 10.1111/j.0013-9580.2004.01304.x.
- G. Battistella et al., ‘Robust thalamic nuclei segmentation method based on local diffusion magnetic resonance properties’, Brain Struct Funct, vol. 222, no. 5, pp. 2203–2216, Jul. 2017, doi: 10.1007/s00429-016-1336-4.
- A. Deistung, A. Schäfer, F. Schweser, U. Biedermann, R. Turner, and J. R. Reichenbach, ‘Toward in vivo histology: A comparison of quantitative susceptibility mapping (QSM) with magnitude-, phase-, and R2⁎-imaging at ultra-high magnetic field strength’, NeuroImage, vol. 65, pp. 299–314, Jan. 2013, doi: 10.1016/j.neuroimage.2012.09.055.
- A. Abosch, E. Yacoub, K. Ugurbil, and N. Harel, ‘An Assessment of Current Brain Targets for Deep Brain Stimulation Surgery With Susceptibility-Weighted Imaging at 7 Tesla’, Neurosurgery, vol. 67, no. 6, pp. 1745–1756, Dec. 2010, doi: 10.1227/NEU.0b013e3181f74105.
- J. P. Marques and R. Gruetter, ‘New Developments and Applications of the MP2RAGE Sequence - Focusing the Contrast and High Spatial Resolution R1 Mapping’, PLoS ONE, vol. 8, no. 7, p. e69294, Jul. 2013, doi: 10.1371/journal.pone.0069294.
- T. Tourdias, M. Saranathan, I. R. Levesque, J. Su, and B. K. Rutt, ‘Visualization of intra-thalamic nuclei with optimized white-matter-nulled MPRAGE at 7T’, NeuroImage, vol. 84, pp. 534–545, Jan. 2014, doi: 10.1016/j.neuroimage.2013.08.069.
- J. Jorge et al., ‘Improved susceptibility‐weighted imaging for high contrast and resolution thalamic nuclei mapping at 7T’, Magn Reson Med, vol. 84, no. 3, pp. 1218–1234, Sep. 2020, doi: 10.1002/mrm.28197.
- J. P. Marques, T. Kober, G. Krueger, W. van der Zwaag, P.-F. Van de Moortele, and R. Gruetter, ‘MP2RAGE, a self bias-field corrected sequence for improved segmentation and T1-mapping at high field’, NeuroImage, vol. 49, no. 2, pp. 1271–1281, Jan. 2010, doi: 10.1016/j.neuroimage.2009.10.002.
- T. Liu, P. Spincemaille, L. de Rochefort, B. Kressler, and Y. Wang, ‘Calculation of susceptibility through multiple orientation sampling (COSMOS): A method for conditioning the inverse problem from measured magnetic field map to susceptibility source image in MRI: COSMOS Susceptibility Imaging’, Magn. Reson. Med., vol. 61, no. 1, pp. 196–204, Jan. 2009, doi: 10.1002/mrm.21828.
- M. Kanowski, J. Voges, L. Buentjen, J. Stadler, H.-J. Heinze, and C. Tempelmann, ‘Direct Visualization of Anatomic Subfields within the Superior Aspect of the Human Lateral Thalamus by MRI at 7T’, American Journal of Neuroradiology, vol. 35, no. 9, pp. 1721–1727, Sep. 2014, doi: 10.3174/ajnr.A3951.
- J. H. Su et al., ‘Thalamus Optimized Multi Atlas Segmentation (THOMAS): fast, fully automated segmentation of thalamic nuclei from structural MRI’, NeuroImage, vol. 194, pp. 272–282, Jul. 2019, doi: 10.1016/j.neuroimage.2019.03.021.
- K. Niemann, V. R. Mennicken, D. Jeanmonod, and A. Morel, ‘The Morel Stereotactic Atlas of the Human Thalamus: Atlas-to-MR Registration of Internally Consistent Canonical Model’, NeuroImage, vol. 12, no. 6, pp. 601–616, Dec. 2000, doi: 10.1006/nimg.2000.0650.
- G. Schaltenbrand and W. Wahren, ‘Atlas for Stereotaxy of the Human Brain’, Chicago: Year Book Medical Publishers, 1977.
Figures
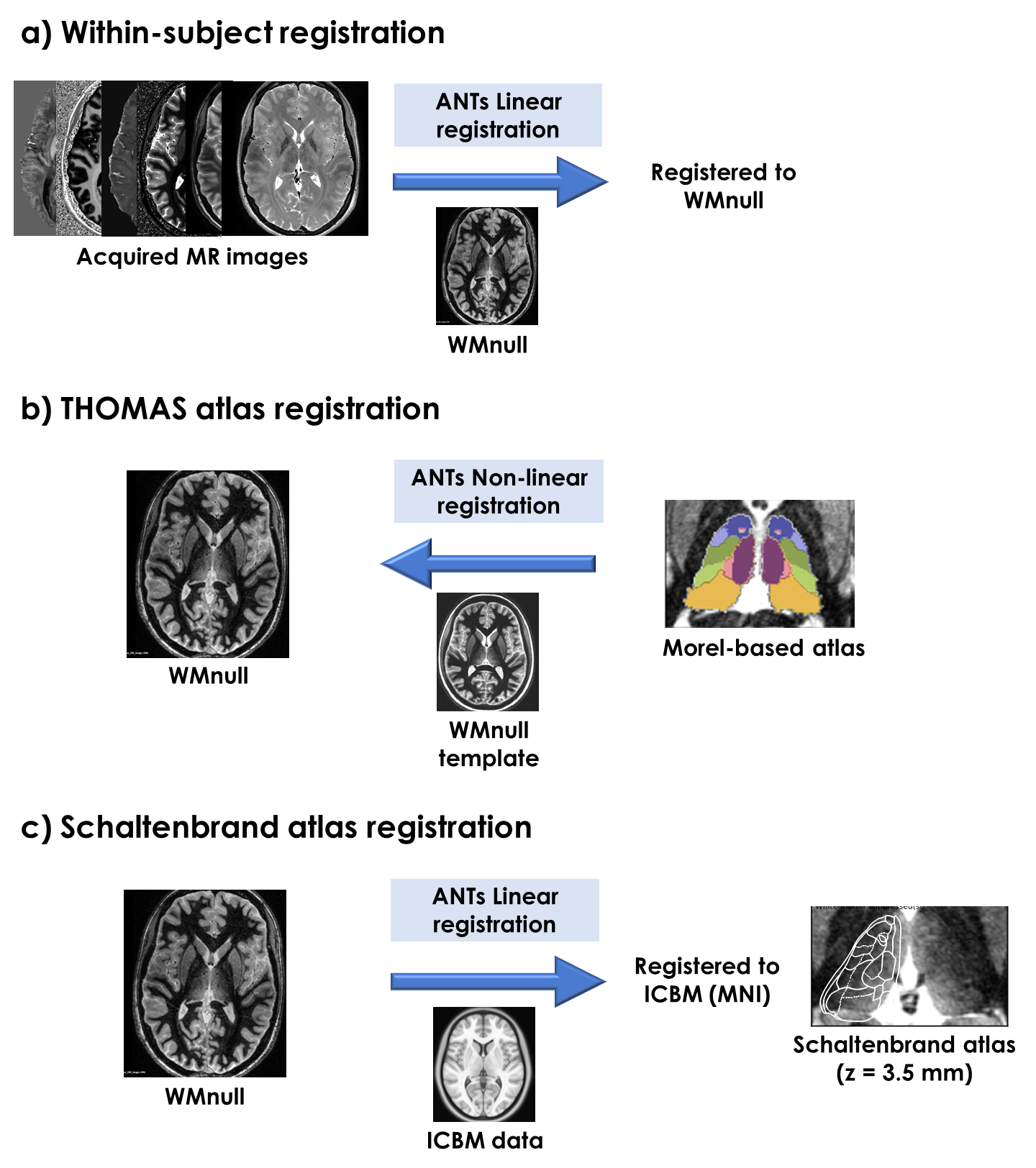
Figure 1. Schematic outline of
data processing. a) Registration of acquired MR images to WMnull, using
ANTs. b) Implementation of the THOMAS approach to fit a Morel-based thalamic
atlas to the WMnull anatomy. c) Registration of the in-vivo data to MNI,
for alignment with the Schaltenbrand atlas.
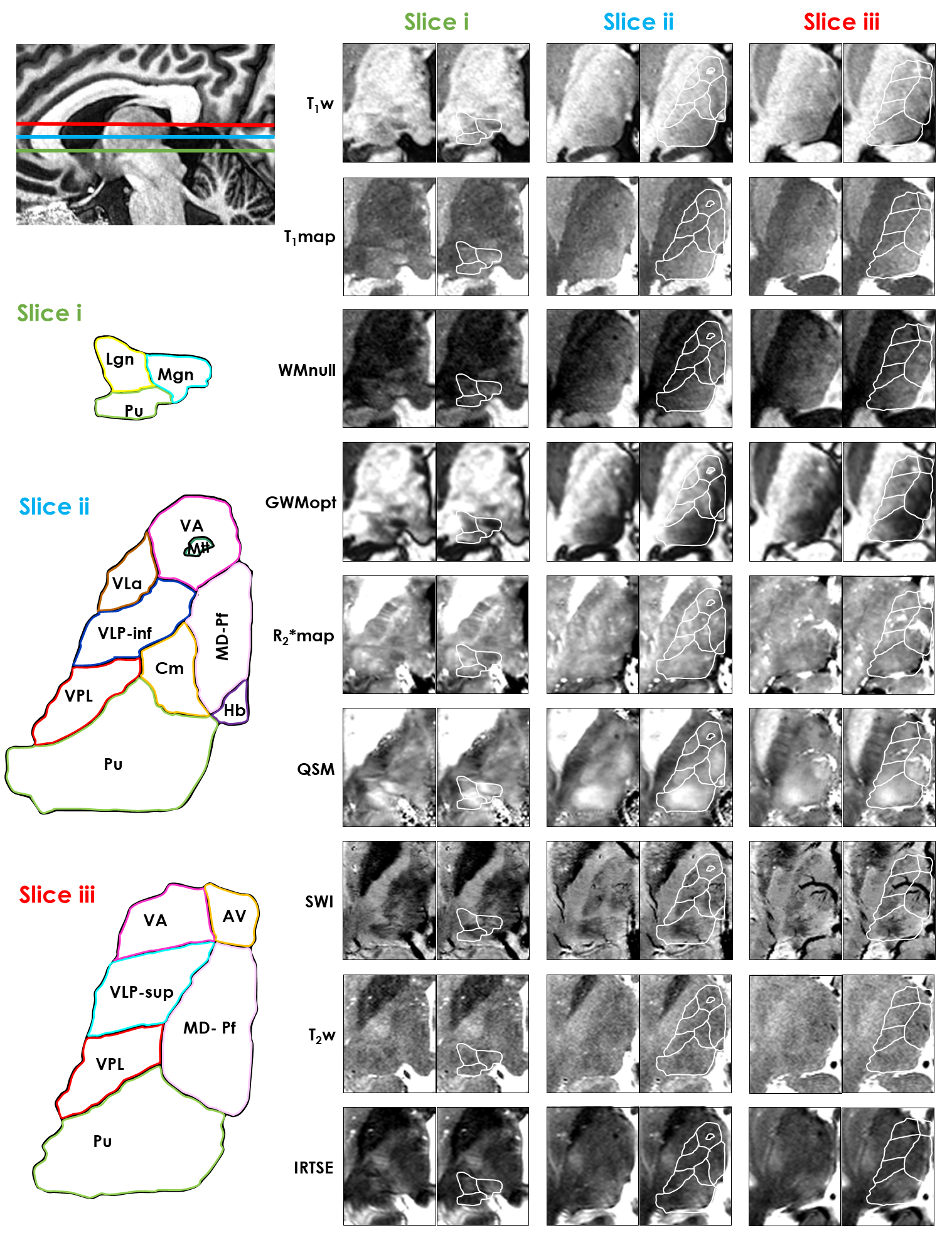
Figure 2. Visualization of
three in-vivo thalamic axial slices with multimodal MRI, compared against the
Morel-based atlas (THOMAS). The anatomical
location of each slice is shown on the upper left corner. The atlas labels are
indicated on the lower left. Each in-vivo image slice (right side) is shown
with and without an atlas overlay.
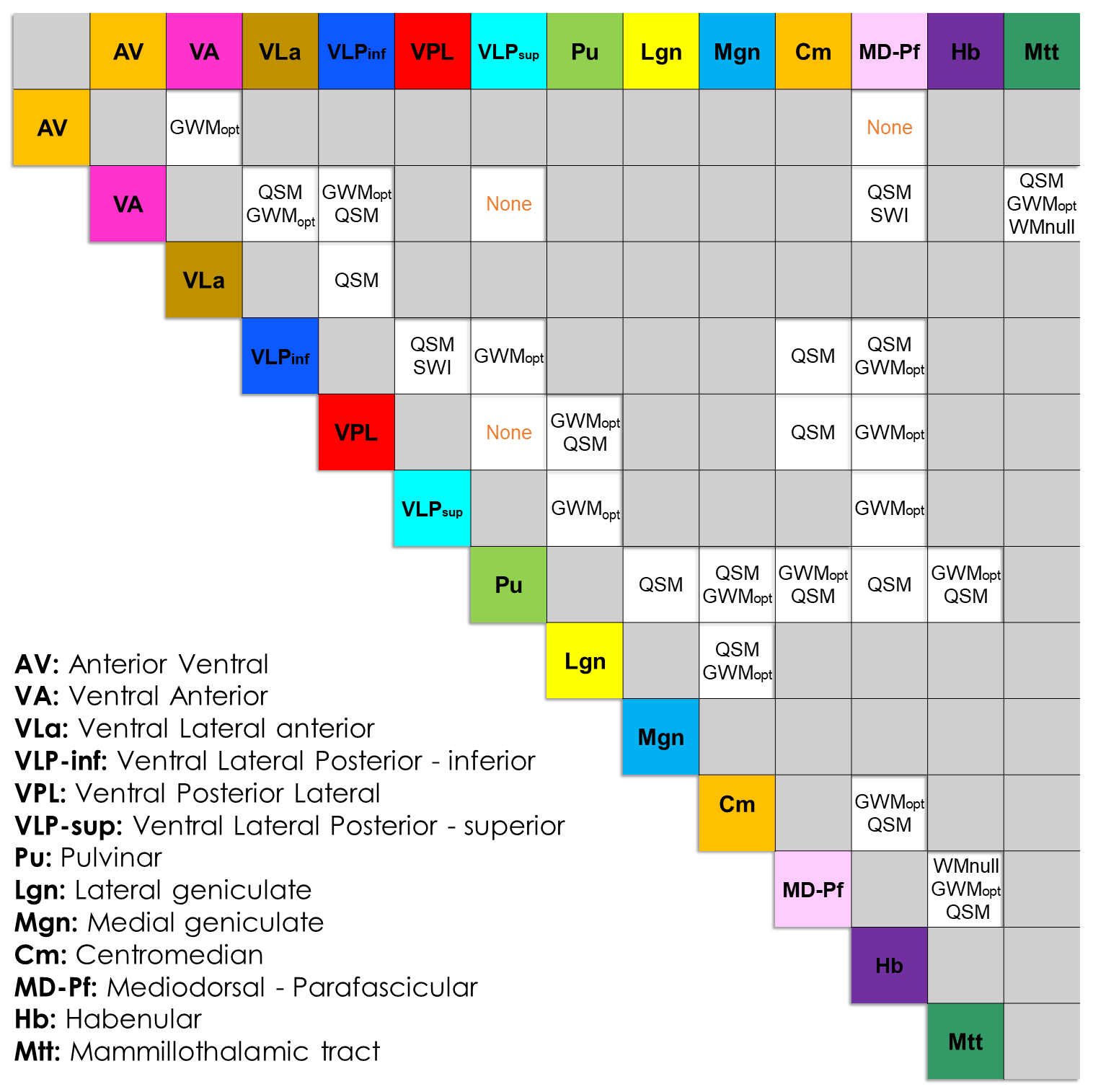
Figure 3. Identification of the
imaging modality/ies that best differentiate each pair of thalamic nuclei, for
the Morel-based atlas. This result represents a consensus between the two
experts; cases, where one or both experts decided that no modality could offer a clear differentiation, are indicated with “None” (orange). Pairs of non-adjacent
nuclei are left empty (gray). The atlas labels are defined on the lower left
corner.
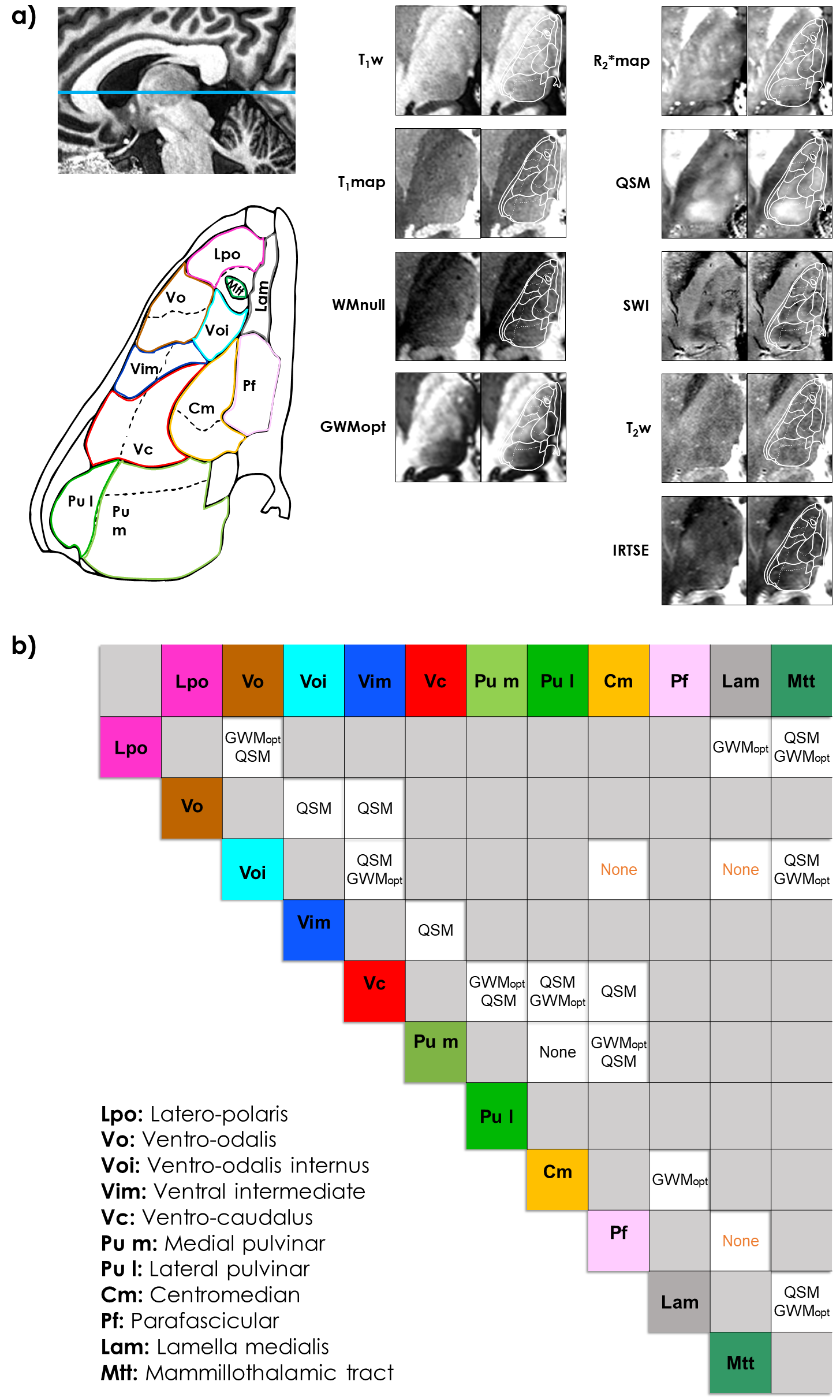
Figure 4. a) Visualization of an in-vivo thalamic axial
slice (MNI z=3.5mm), compared with the Schaltenbrand atlas. The slice location
is shown on the upper left, and atlas labels on the lower left. Each in-vivo
slice (right side) is shown with and without overlay. b) Identification
of the modality/ies that best differentiate each pair of nuclei, as a consensus
between the two experts. Cases, where one or both experts decided that no modality could offer a clear differentiation, are indicated with “None”
(orange). Non-adjacent nuclei are in gray. Labels are defined on the lower
left.
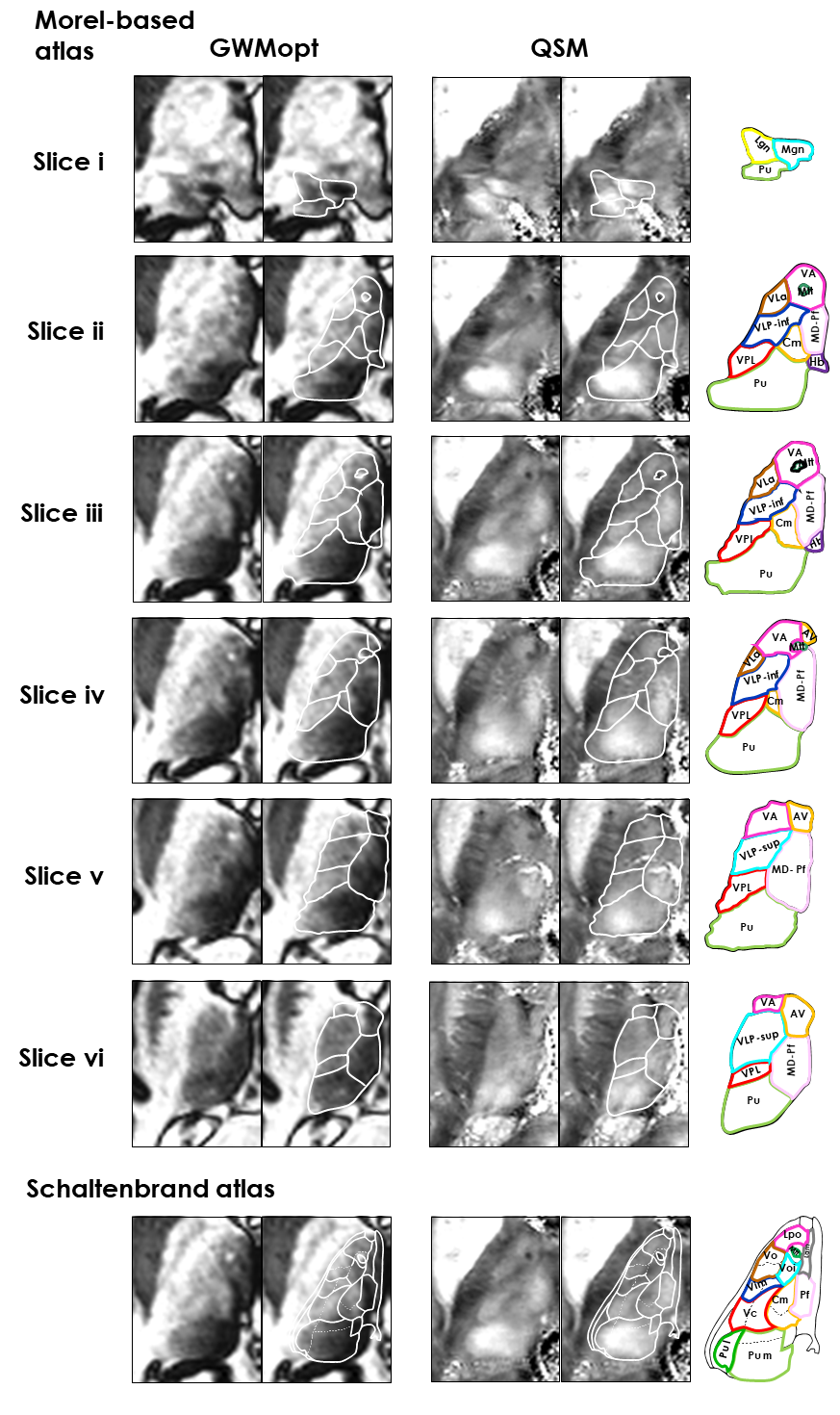
Figure 5. Expanded visualization of
thalamic slices for the two most valuable modalities identified in this work:
QSM and GWMopt. The two modalities are shown in comparison with the Morel-based
atlas (THOMAS; six slices) and Schaltenbrand atlas (z=3.5mm). The Morel slices
i, iii and v correspond to slices i–iii of Fig.2.
DOI: https://doi.org/10.58530/2023/1357