3775
Developmental pattern of association fibers and their interaction with associated cortical microstructures in 0-5 month-old infants1Key Laboratory for Biomedical Engineering of Ministry of Education, Department of Biomedical Engineering, College of Biomedical Engineering & Instrument Science, Zhejiang University, Hangzhou, China, 2Department of Radiology, Children's Hospital, Zhejiang University School of Medicine, Hangzhou, China, 3Department of Child Health, Children's Hospital, Zhejiang University School of Medicine, Hangzhou, China
Synopsis
The present study aimed to investigate the spatiotemporal developmental pattern of association fibers in infants aged 0-5 months and the interaction between these fibers and the associated cortex utilizing FBA based on HARDI data. We found that the C-shaped fibers demonstrate an approximately symmetrical along-track pattern with more advanced development in the middle segments than the extremities; the horizontally oriented fibers manifest that the anterior segments started later but developed faster than the posterior segments. Mediation analysis revealed the mediation effect of cortical GM on the development of WM was more prominent than that of WM on GM.
Introduction
Association fibers connect the ipsilateral cortex and are involved in cognitive and emotional functions1, and experience rapid myelination during infancy2. Yet, the spatiotemporal patterns of microstructural changes along these tracts across time remain largely unknown. In addition, despite the close relationship between WM and cortical GM, how they interact with each other, e.g, whether WM plays a role in shaping the cortical GM development or vise versa, has not been addressed in previous studies. In this study, we aimed to 1) first characterize the along-tract developmental pattern of major association fibers in infants aged 0-5 months using fixel-based analysis (FBA)3,4 on HARDI data; and 2) to elucidate the interaction between these association fibers and their associated cortical GM using mediation analysis.Methods
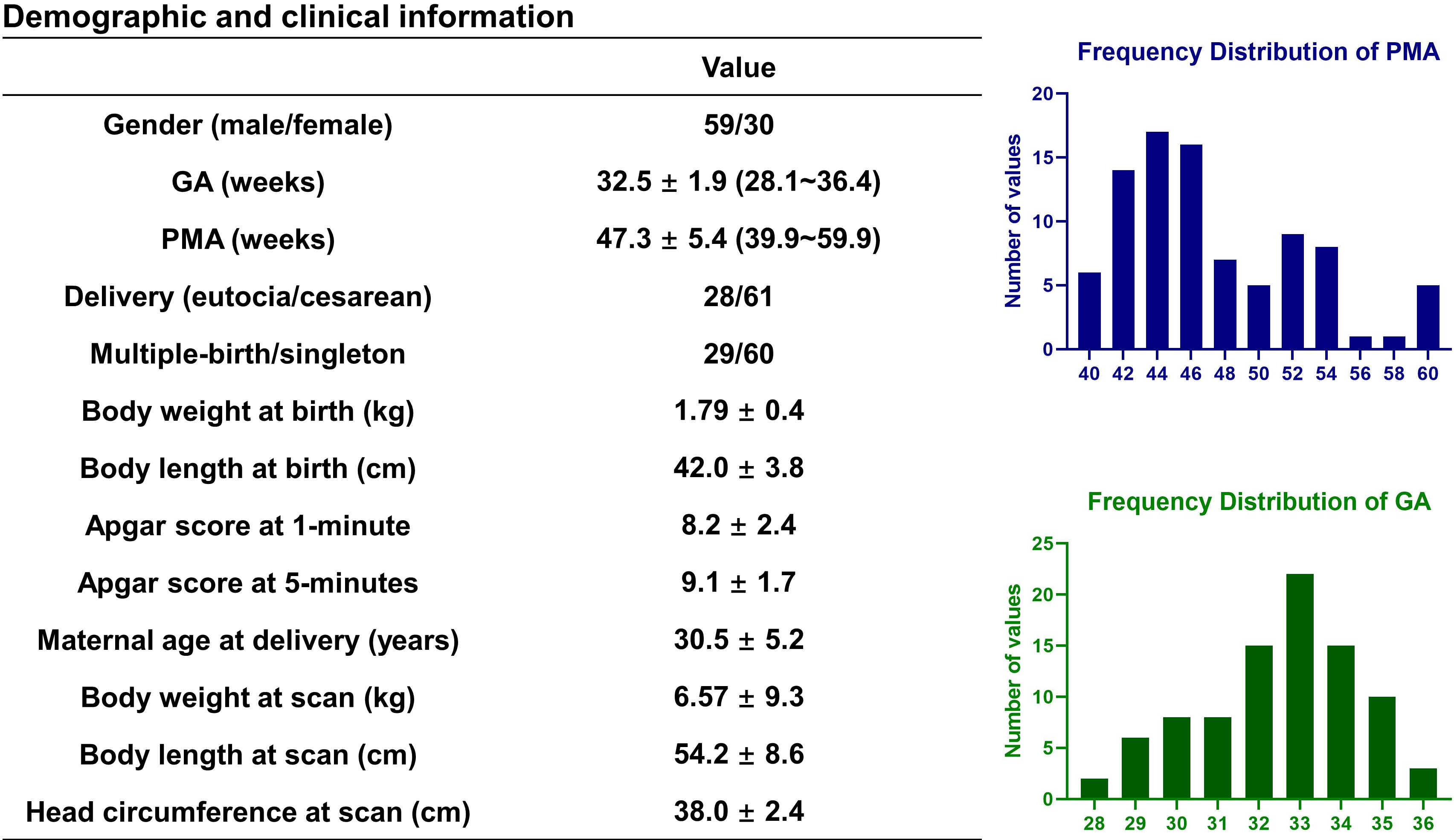
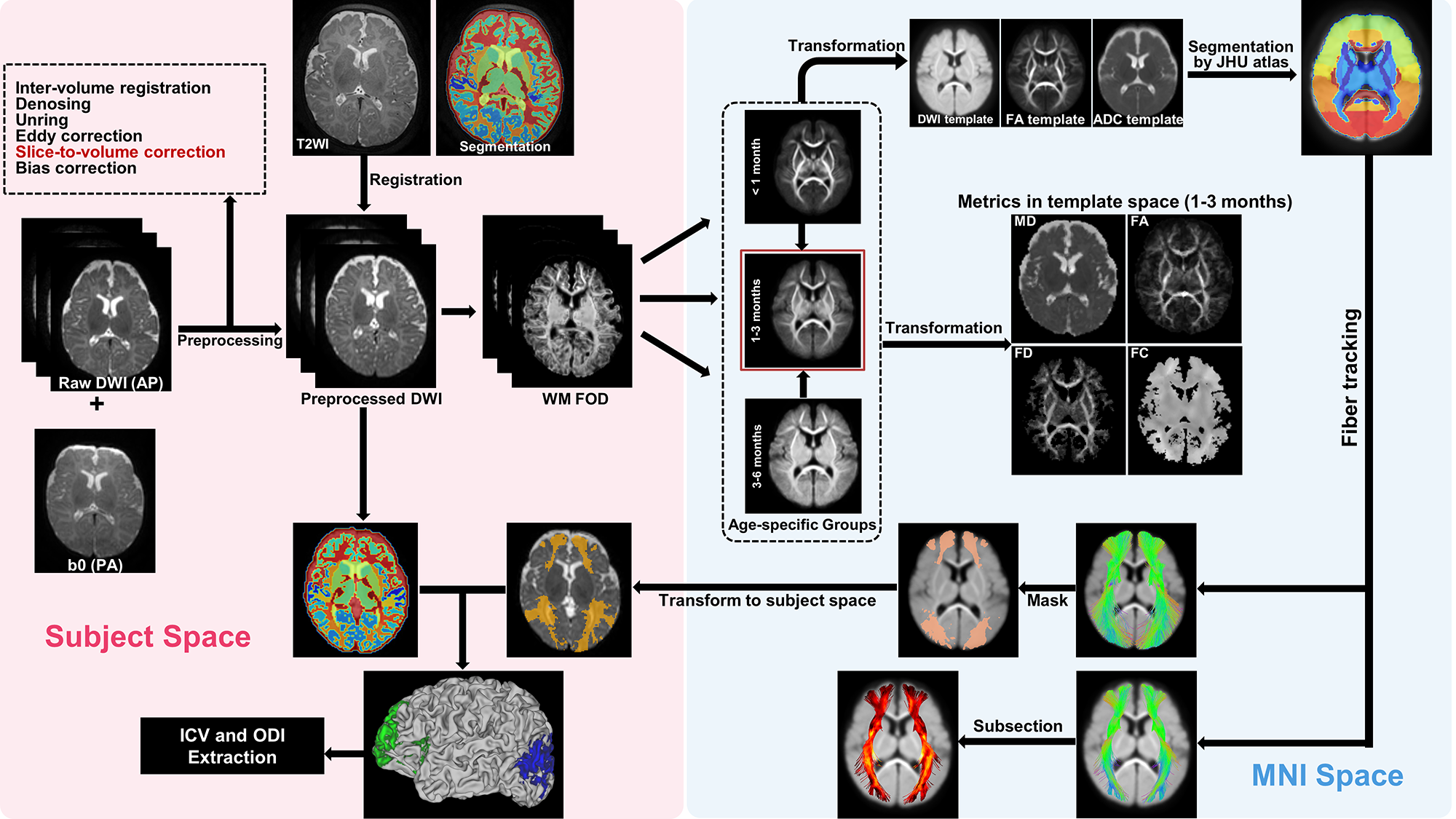
Data acquisition: Eighty-nine healthy preterm-born infants with gestational age (GA) at birth between 28.1~36.4 weeks and postmenstrual age (PMA) at scan between 39.9~59.9 weeks (Fig.1). MRI was performed on a Philips 3.0T Achieva system, including 1) 3D T2-weighted image: TR/TE=2200/278 ms, flip angle=90°, voxel size=0.75×0.75×0.8 mm3, FOV=180×180 mm2; 2) Multi-shell HARDI data acquired with 32 noncolinear diffusion directions and two b-values of 800/1500 s/mm2, one pair of b0s with inverse phase-encoding directions, TR/TE=9652/115 ms, voxel resolution=1.5×1.5×2 mm3, FOV=180×180 mm2.Data processing: The T2-weighted image was segmented into 87 regions using DrawEM5, followed by manual correction of the cortical labels. The preprocessed HARDI data were processed using MRtrix3 MSMT-CSD pipeline6. Participants were categorized into three groups based on PMA. For each group, a specific fiber orientation distribution (FOD) template was generated, then all individual data was transformed to the 1-3 months template using a two-step registration with the age-specific FOD template as an intermediate reference. Next, the FOD template was segmented using the JHU Neonate Atlas7 to track 5 association fibers, and the tracts were divided into 100 segments. FA, MD, fiber density (FD), and fiber cross-section (FC) indices were obtained from each segment. Finally, the cortical GM regions connected to the association tracts were extracted as cortical voxels containing the fiber bundles. Based on the NODDI model, the intracellular volume (ICV) and orientation dispersion index (ODI) of GM associated with the anterior and posterior ends of the fibers were obtained8 (Fig.2).
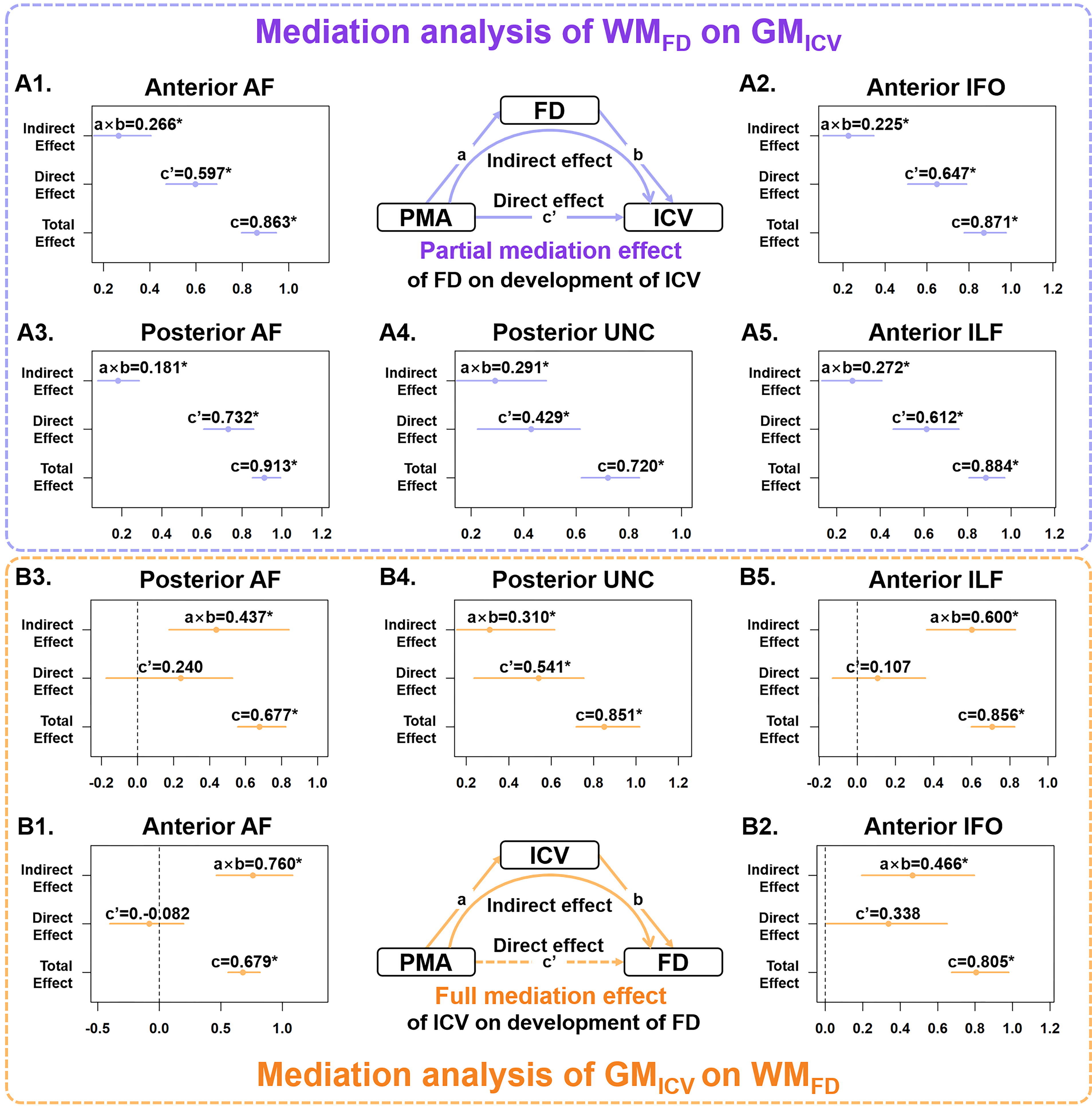
Statistical analysis: Mediation analysis9 was performed for detecting the mediation effect of WM (FD and FC) on the PMA and GM (ICV and ODI), and also that of GM on the PMA and WM (Fig.5), with gender, birthweight at birth, and GA as covariates.
Results
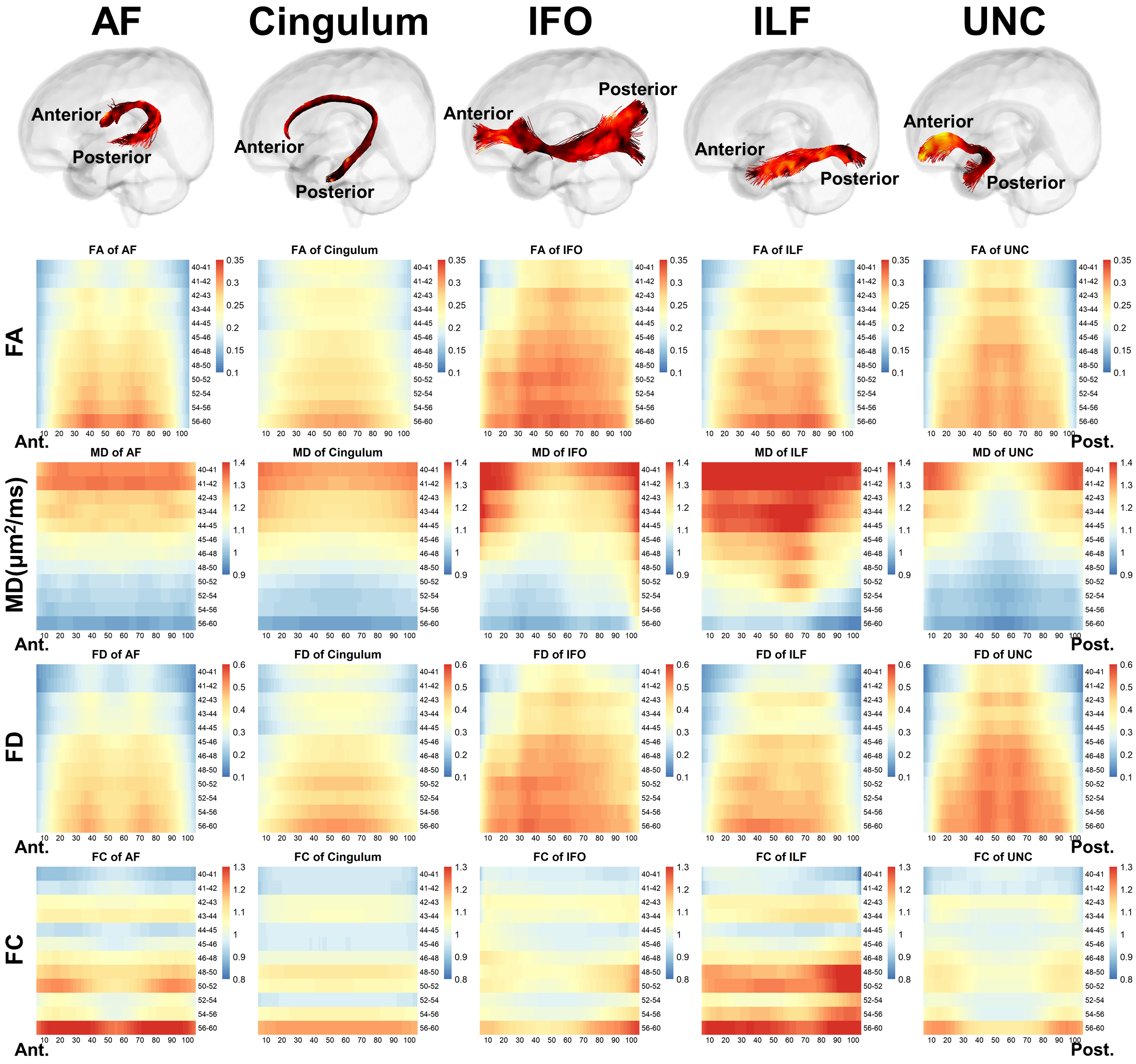
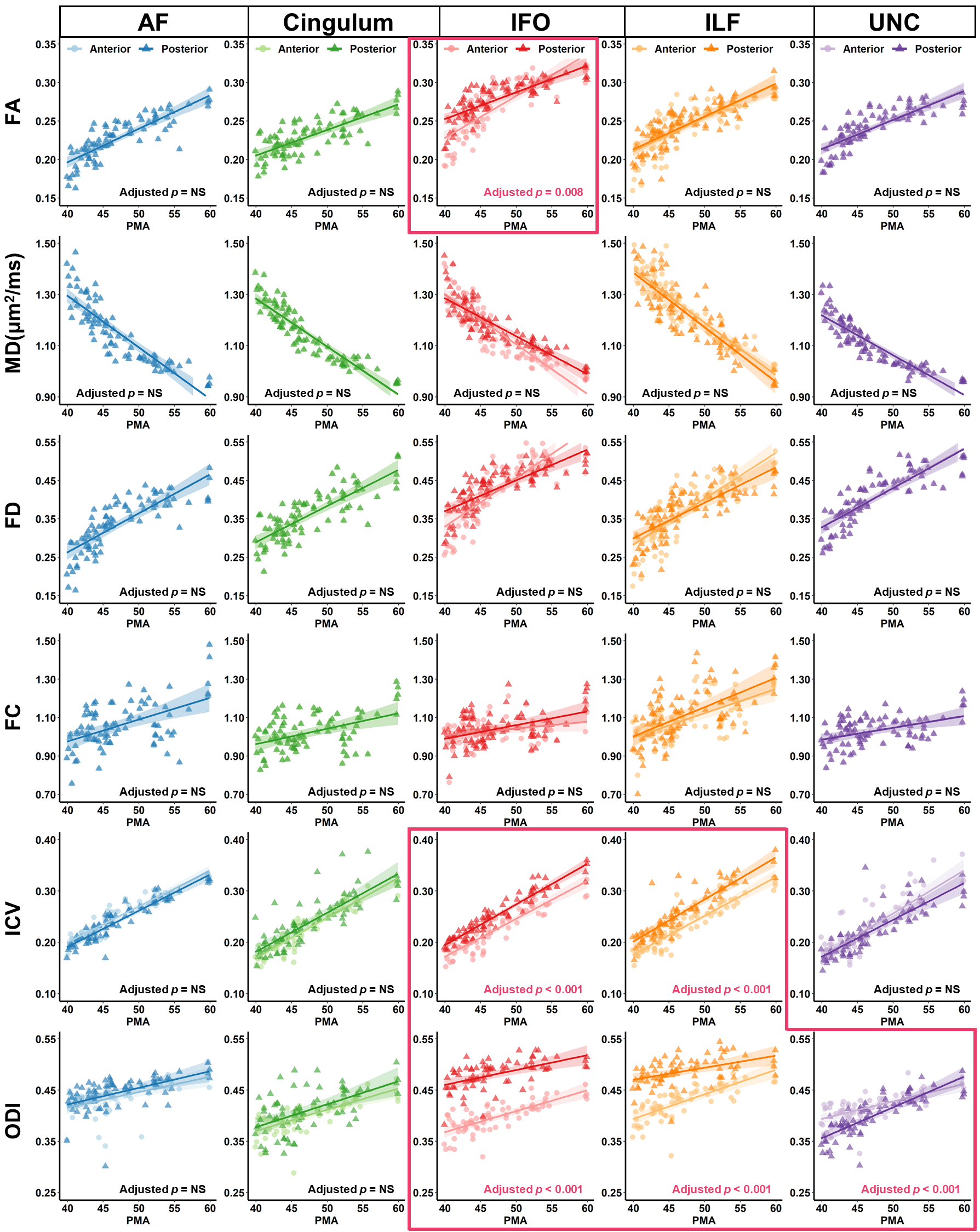
Distinct spatiotemporal developmental patterns of five association fibers were demonstrated in Fig.3. We found that C-shaped fibers, including arcuate fasciculus (AF), cingulum, and uncinate fasciculus (UNC) demonstrate symmetrical along-tract patterns with more advanced development in the middle segments than the extremities; and the horizontally oriented fibers, including the inferior fronto-occipital fasciculus (IFO) and inferior longitudinal fasciculus (ILF), manifest that the anterior segment started later but developed faster than the posterior segments, e.g., FA of IFO was lower in the anterior segments during 40-52 weeks but exceeded the posterior after 52 weeks (adjusted p = 0.008) (Fig.4). We found the ICV and ODI in cortical GM attached to the posterior end of IFO and ILF continued to be significantly higher than that of the anterior end during 0-5 months (Fig.4).Mediation analysis revealed that except for the cingulum, all other fibers demonstrated a developmental interaction of WM and GM, and the effect of GM on the development of WM is more prominent than the effect of WM on GM (Fig.5). For anterior and posterior AF, anterior IFO, and anterior ILF, a partial mediation effect of FD was found on the development of ICV, manifested as both direct effect of PMA and indirect effect mediated by FD on ICV were significant (Fig.5A). And a full mediation effect of ICV on the development of FD was found, demonstrated as a significant indirect effect of ICV on the development of FD, but a nonsignificant direct effect of PMA on FD (Fig.5B). For all association fibers, no indirect effect associated with the occipital lobe (posterior IFO and ILF) was found, while a developmental interaction between WM and GM associated with the frontal lobe (anterior AF) and temporal lobe (posterior AF and UNC) were significant.
Discussion and Conclusion
Distinct along-tract developmental trajectories of five association fibers were characterized, and the spatiotemporal pattern may reflect the caudal-rostral, central-peripheral developmental order of the infant's brain. The developmental interaction of WM and GM was revealed by the mediation analysis using FBA and NODDI-based microstructural parameters, which revealed a lopsided interaction between GM and WM, and between the anterior and posterior parts of the tracts. The ICV in GM was found to have a full mediation effect on the FD of WM, which indicates the axonal packing reflected by the FD was mainly influenced by the neurite density reflected by ICV10,11. A partial mediation effect of FD was found on the ICV, possibly due to the packing density influences the signal propagation12, and therefore make a certain impact on cortical maturation.Acknowledgements
This work was supported by the Ministry of Science and Technology of the People’s Republic of China (2018YFE0114600), National Natural Science Foundation of China (61801424, 81971606, 82122032), and Science and Technology Department of Zhejiang Province (202006140).References
1. Philippi CL, Mehta S, Grabowski T, Adolphs R, Rudrauf D. Damage to Association Fiber Tracts Impairs Recognition of the Facial Expression of Emotion. J Neurosci. 2009;29(48):15089-15099.
2. Dubois J, Dehaene-Lambertz G, Kulikova S, Poupon C, Huppi PS, Hertz-Pannier L. The Early Development of Brain White Matter: A Review of Imaging Studies in Fetuses, Newborns and Infants. Neuroscience. 2014;276:48-71.
3. Raffelt DA, Smith RE, Ridgway GR, et al. Connectivity-based fixel enhancement: Whole-brain statistical analysis of diffusion MRI measures in the presence of crossing fibres. Neuroimage. 2015;117:40-55.
4. Raffelt DA, Tournier JD, Smith RE, et al. Investigating white matter fibre density and morphology using fixel-based analysis. Neuroimage. 2017;144:58-73.
5. Makropoulos A, Gousias IS, Ledig C, et al. Automatic Whole Brain MRI Segmentation of the Developing Neonatal Brain. Ieee T Med Imaging. 2014;33(9):1818-1831.
6. Jeurissen B, Tournier JD, Dhollander T, Connelly A, Sijbers J. Multi-tissue constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI data. Neuroimage. 2014;103:411-426.
7. Oishi K, Mori S, Donohue PK, et al. Multi-contrast human neonatal brain atlas: application to normal neonate development analysis. Neuroimage. 2011;56(1):8-20.
8. Zhang H, Schneider T, Wheeler-Kingshott CA, Alexander DC. NODDI: Practical in vivo neurite orientation dispersion and density imaging of the human brain. Neuroimage. 2012;61(4):1000-1016.
9. Tingley D, Yamamoto T, Hirose K, Keele L, Imai K. mediation: R Package for Causal Mediation Analysis. J Stat Softw. 2014;59(5).
10. Bandeira F, Lent R, Herculano-Houzel S. Changing numbers of neuronal and non-neuronal cells underlie postnatal brain growth in the rat. P Natl Acad Sci USA. 2009;106(33):14108-14113.
11. van Dyck LI, Morrow EM. Genetic control of postnatal human brain growth. Curr Opin Neurol. 2017;30(1):114-124.
12. Rama S, Zbili M, Debanne D. Signal propagation along the axon. Curr Opin Neurobiol. 2018;51:37-44.
Figures