2539
Suppression of extracranial lipids in short TE STEAM of the human brain at 7T with an external crusher coil1Radiology, University Medical Center Utrecht, Utrecht, Netherlands
Synopsis
Short TE STEAM is an appealing MRS sequence especially for the quantification of low-concentration J-coupled spin systems. However, in comparison to sLASER, STEAM is associated with larger chemical shift displacement errors in all three directions. With an inhomogeneous B1+-field, slice profiles degrade, resulting in artefacts from extracranial lipids. Here we demonstrate that lipid artefacts can be removed through the use of an external crusher coil. Without use of the crusher coil, large extracranial lipid artefacts are present when selecting a large voxel. Upon activation of the crusher coil, these signals are eliminated, yielding 1H spectra free of large baseline distortions.
Introduction
MRS of the human brain at 7T enables the identification of a large number of metabolites, with increased spectral resolution and SNR compared to lower field strengths. Typically, artifacts associated with lipid signal contamination from extracranial sources are an issue when performing spectroscopic imaging, but lipids can also be a problem in single-voxel acquisitions. Particularly in cases where the voxel size must be increased for sufficient SNR to detect low-concentration metabolites such as glutamine and glutathione (GSH). Here sequences with short echo time (TE) are beneficial as they reduce T2-mediated signal loss, limit the J-coupling evolution for coupled spin systems, and are generally assumed to reduce quantification errors1.The STEAM sequence is suited for acquiring short TE spectra but, when using volume transmitters with typically a lower B1+, it suffers from large chemical shift displacement errors, which worsen with increasing field strength. Moreover, the slice profile of the radio-frequency pulses (RF) used for localization depends on the B1+ field and may excite extracranial lipids outside the voxel of interest, where B1+ levels can substantially deviate, leading to contamination.
One possible solution is the use of outer-volume suppression pulses; however, these will increase overall power-deposition (and thus repetition time (TR)), require slice by slice B1+ calibration and suffer from their own limitations with respect to spatial profile. To address the issue of lipid contamination for short TE single-voxel MRS, we propose the use of a crusher coil generating an external suppression field2.
Methods
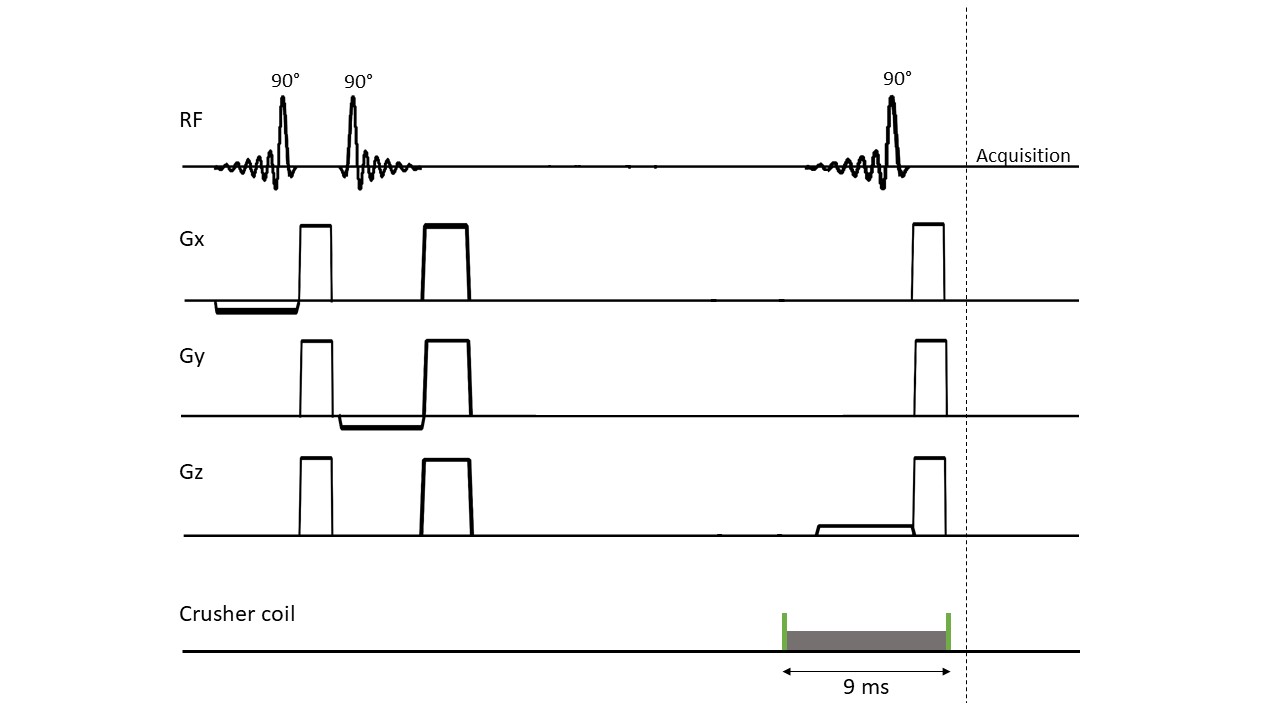
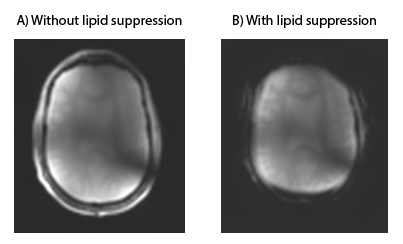
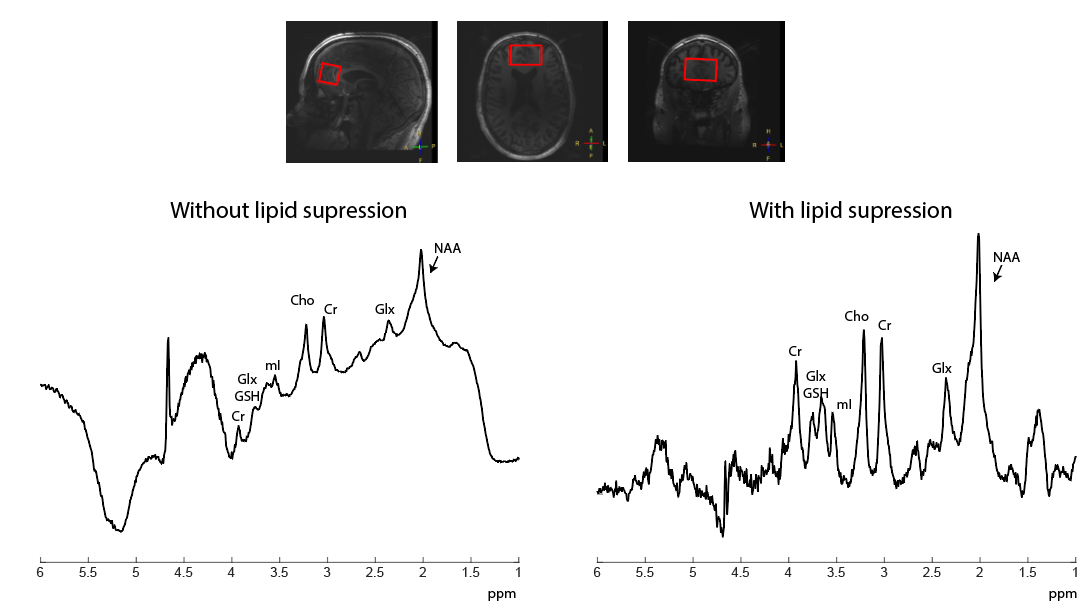
A 7T MRI scanner (Philips, Best, the Netherlands) was used, equipped with eight transmit/receive 1H antennas, combined with a 32-channel receive array (Nova Medical, Wilmington, United States). STEAM 1H MRS data (TE 7.8 ms, TM 30 ms, TR 3000 ms, NSA 32, VAPOR 300 Hz, voxel size 30x50x30 mm3) was acquired in a healthy volunteer (male, 47 years). B1+ and B0 shimming were performed on a manually drawn region of interest (ROI) in the frontal lobe. The STEAM voxel was planned in a region containing the pre-frontal cortex and care was taken not to include any extracranial lipids.Lipid suppression was performed using an external coil that was placed inside the 32-channel receive coil. This coil was driven by the Z3 (third order) shim amplifier, controlled using a dynamic shim updating unit (DSU: RRI, Billerica, USA ). Two TTL pulses (6 ms before and 3 ms after the top of the last STEAM pulse) were added to the STEAM sequence to activate and deactivate the crusher coil, respectively. The crusher coil pulsed with a 3A current over 9 ms per TR (Figure 1). Lipid suppression was checked qualitatively by applying comparable crushing fields during the acquisition of a 2D FFE image (Figure 2). STEAM spectra were recorded from the same voxel with and without activating the crusher coil.
Results
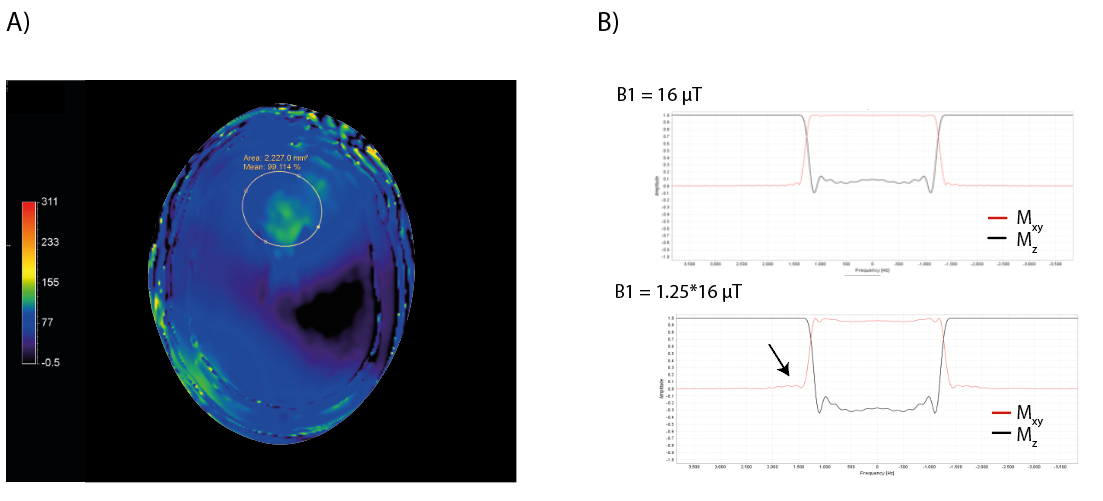
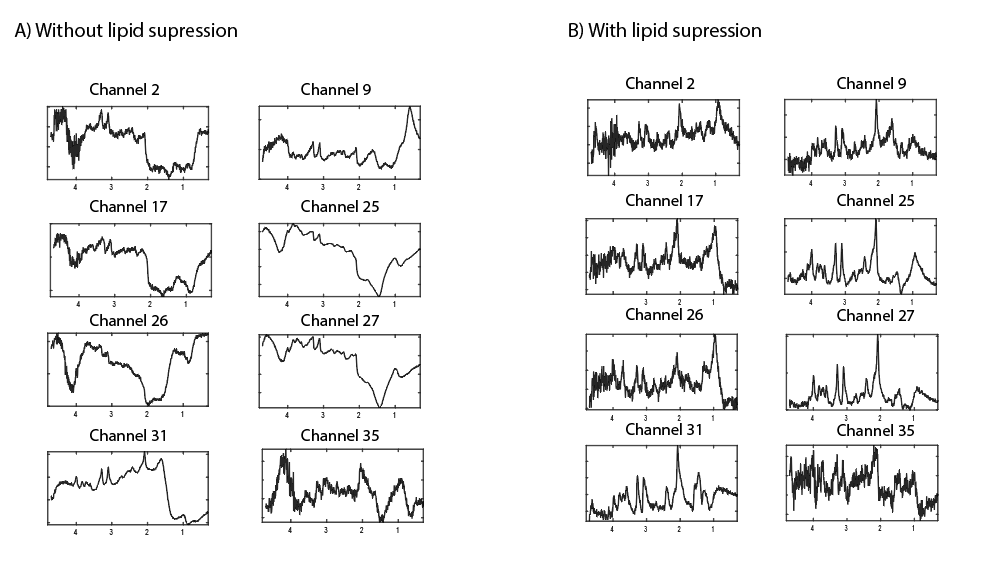
Figure 3 shows the B1+ map after B1+ shimming, clearly indicating substantial differences in B1+ inside and outside (e.g. at the skull) the shimmed ROI in the frontal lobe, which can affects the slice profile. Figure 4 shows 1H STEAM spectra for several receive channels without and with the use of the crusher coil. Extracranial lipid artefacts can be found in many of the receive channels without crushing, while they are greatly reduced with crushing. Figure 5 shows the channel-combined spectra without and with the use of the crusher coil. Without crushing, large extracranial lipid artefacts are present, making reliable signal quantification impossible. Upon lipid suppression with the crusher coil, the lipid signals are to a great extent eliminated, yielding a spectrum free of large baseline distortions.Discussion and Conclusion
Short TE single-voxel 1H MRS at 7T is appealing especially for the quantification of low-concentration, J-coupled spin systems. Unfortunately these sequences are susceptible to corruption from lipid artefacts. In this work, we demonstrate that lipid artefacts can be removed through the use of an external suppression field generated by a crusher coil.Acknowledgements
No acknowledgement found.References
1. Wilson, M, Andronesi, O, Barker, PB, et al. Methodological consensus on clinical proton MRS of the brain: Review and recommendations. Magnetic Resonance in Medicine. 2019; 82, 527–550
2. Boer, VO, Van De Lindt, T, Luijten, PR & Klomp, DWJ. Lipid suppression for brain MRI and MRSI by means of a dedicated crusher coil. Magn. Reson. Med. 2015; 73, 2062–2068
Figures