1645
In-vivo microstructural characterisation of first and higher order thalamic nuclei: a diffusion kurtosis imaging study1CIMeC, Center for Mind/Brain Sciences, University of Trento, Rovereto (Trento), Italy
Synopsis
The thalamus is a primary station for information processing within the brain. It is composed of a collection of nuclei with different histological and functional properties, classified as First Order (FO, processing sensory input) or Higher Order (HO, playing associative roles). We used diffusion-weighted MRI (dMRI) to investigate microstructural properties of FO and HO thalamic nuclei, focusing on fractional anisotropy, mean diffusivity, and mean kurtosis derived from Diffusion Kurtosis Imaging (DKI). This initial evidence suggests that DKI might capture distinctive features of FO and HO thalamic nuclei, and paves the way to further studies investigating nucleus-specific microstructural effects with DKI.
Introduction
The thalamus is a collection of grey matter nuclei primarily involved in gating ascending cortical input and modulating cortical activity via transthalamic cortico-cortical pathways1. Histological research2,3 has differentiated thalamic nuclei in terms of microstructural features such as myelination, cell size and cell density. A more functional classification distinguishes thalamic nuclei as supporting First Order (FO) or Higher Order (HO) functions4. Despite this, the thalamus is often considered a uniform structure in MRI studies, except for the recent possibility of segmenting its nuclei from T1 anatomical MRI5. However, it is yet unclear whether diffusion-weighted MRI (dMRI) can capture the microstructural heterogeneity across nuclei. In this study, we evaluate the sensitivity of diffusion MRI to differentiate microstructural properties of thalamic nuclei, in particular to differentiate FO and HO nuclei in terms of DKI-derived fractional anisotropy (FA), mean diffusivity (MD), and mean kurtosis (MK)6,7.Methods
Acquisition: Healthy adults (N=10, mean age (years, sd)=26.7 (4.4)) underwent scanning using a 3T scanner (MAGNETOM Prisma, Siemens Healthcare, Erlangen, Germany) with a 64-channel head-neck RF receive coil, to acquire 3D T1-weighted (T1w, multi-echo-MPRAGE, 1mm-isotropic), and dMRI (2mm-isotropic, TE/TR=76/4200ms, shells: b={0,700,1000,2850} s/mm2, 32/64/64 directions) data.Image processing: T1w images were skull stripped and processed in FreeSurfer v.7.1.1 recon-all. Thalamic nuclei were segmented from T1w images with FreeSurfer’s thalamic segmentation algorithm5 and volumetric data for each nucleus was obtained. Thalamic nuclei larger than 100 mm3 were considered. FO nuclei: medial geniculate (MGN), lateral geniculate (LGN), ventral lateral anterior (VLa), ventral lateral posterior (VLp), ventral posterolateral (VPL). HO nuclei: centromedian (CM), mediodorsal lateral (MDl), mediodorsal medial (MDm), anterior (PuA), inferior (PuI), lateral (PuL), and medial (PuM) divisions of the pulvinar, and ventral anterior (VA). dMRI data were denoised8, Gibbs-ringing corrected9, and corrected for eddy currents, head motion, and geometric distortions in FSL’s eddy v.6.0.310, and for bias field in MRtrix v.3.0.2. dMRI data was then fitted with DKI and FA, MD, and MK maps were derived7. Linear registrations between each participant’s first b=0 volume and T1w image were computed in ANTs and applied to thalamic segmentations in order to extract the median of each DKI-derived metrics from each nucleus segmentation aligned in the original dMRI image space.
Statistical analyses: The Kruskal-Wallis test was used in R11 to investigate the effect of nucleus for each DKI metric. Independent samples t-tests were used in R to assess differences in DKI indices between FO and HO nuclei. All p-values were Bonferroni corrected for the three DKI metrics investigated. Cohen’s d was used to determine the effect size of differences between FO and HO nuclei for each DKI index.
Results
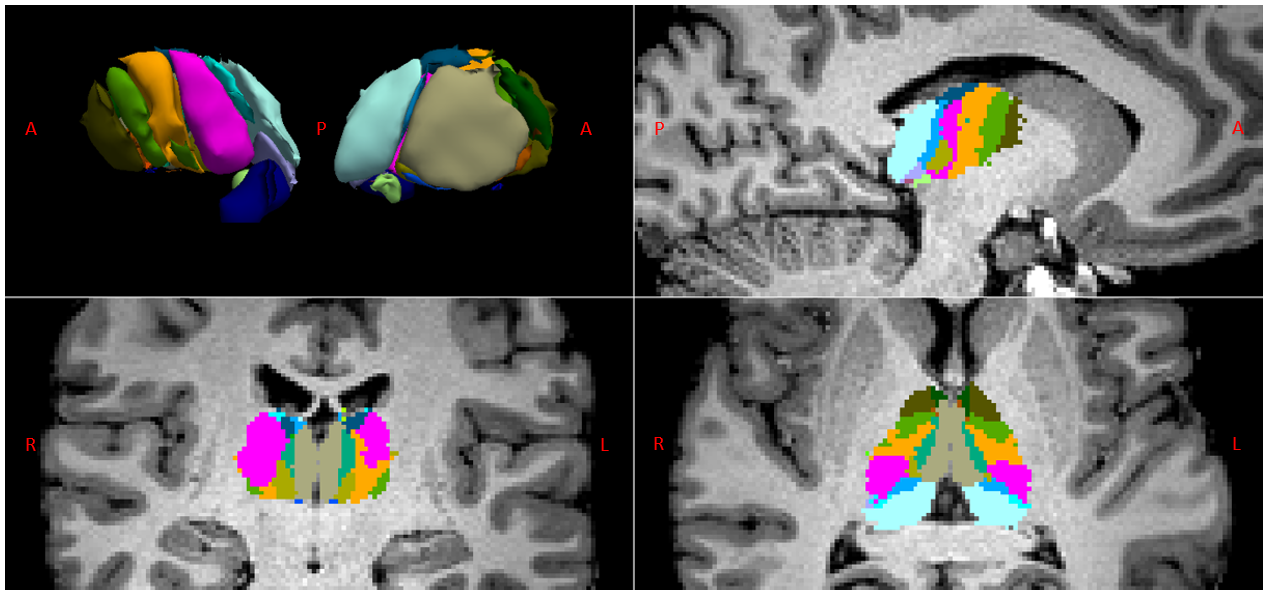
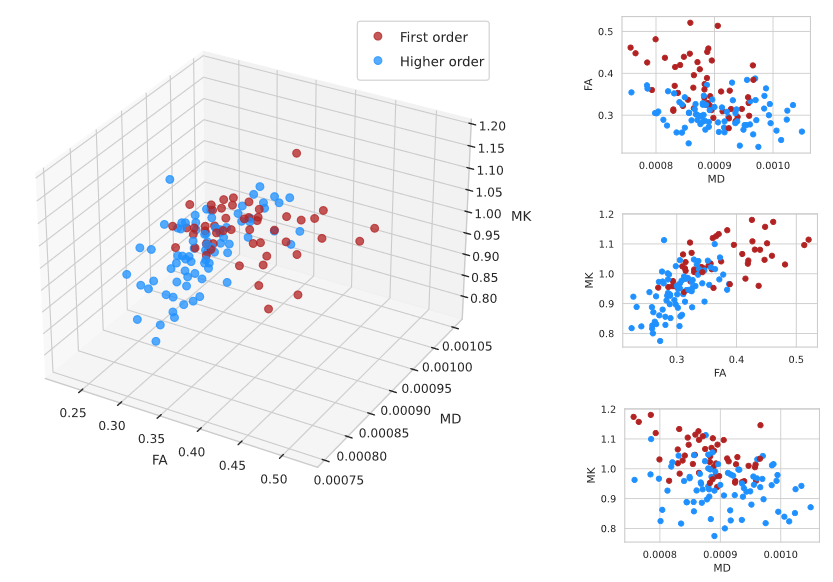
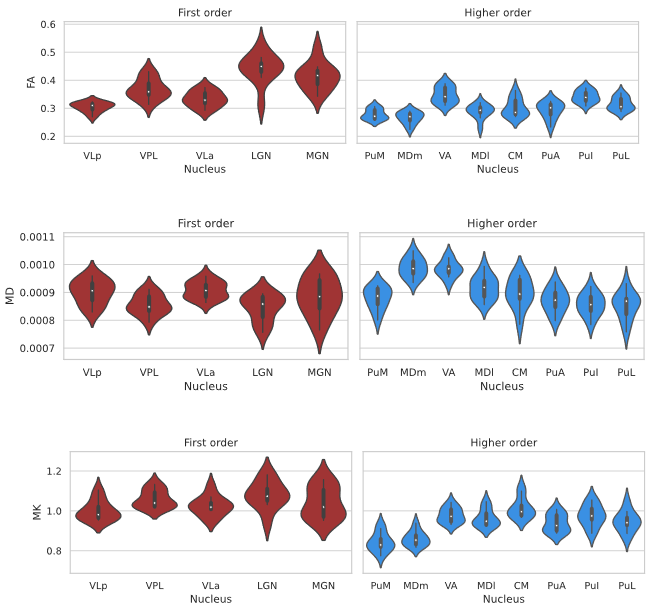
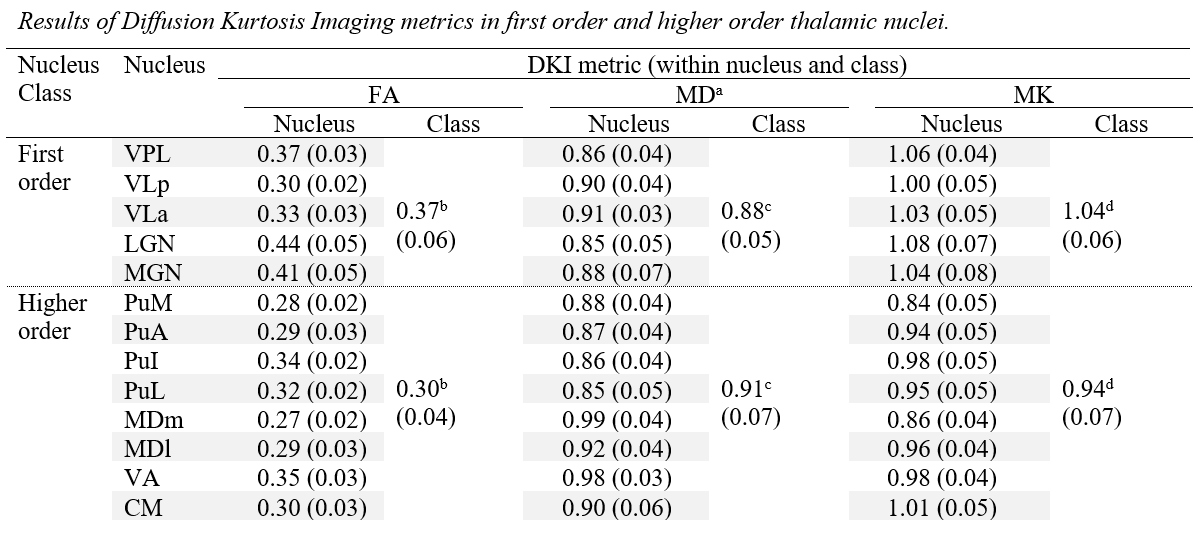
Fig. 1 shows the thalamic parcellations of a representative subject. Volumetric results agreed with the literature5,12. Scatter plots and distributions of median dMRI metrics for the considered nuclei are shown in Fig. 2 and Fig. 3, respectively. Mean (SD) values for all dMRI metrics and nuclei are listed in Fig. 4. Kruskall-Wallis tests detected a significant effect of nucleus for all investigated metrics (FA: χ²=88.17; MD: χ²=61.34; MK: χ²=83.4; all padj<0.0001). Post-hoc t-tests revealed larger FA and MK for FO as compared to HO nuclei (FA: t=6.92; MK: t=8.37, all padj<0.0001), and lower MD in FO than in HO nuclei (t=-2.59, padj<0.05). Both MK and FA showed large effect sizes, with the largest value for MK (dMK=1.47, dFA=1.40), while MD showed a small effect size (dMD=-0.44).Discussion and Conclusions
While the thalamus is often considered as a single structure in MRI studies, our findings indicate that dMRI can capture microstructure heterogeneity associated with thalamic nuclear subdivisions, as expected from histological studies2,3. FA and MK provided higher sensitivity than MD to differentiate between FO and HO nuclei. A DKI-based characterisation of thalamic nuclei has already been attempted in an animal study13, and provided to be in good agreement with histology. Our findings suggest that the combination of DKI metrics such as FA, MD, and MK may help characterise thalamic heterogeneity, and possibly differentiate between FO and HO thalamic nuclei. Since both FA14,15 and MK16,17,18 are associated with restrictions from cellular membranes and myelination, we speculate that the biological underpinnings of such differences in DKI-derived metrics might be represented by different densities of myelinated axonal fibers across the two groups of nuclei. Intriguingly, the potentially larger microstructural complexity of sensory nuclei might reflect the high degree of precision required in these nuclei for sensory processing to be effective19. In conclusion, these findings suggest that DKI metrics may be used to study microstructural changes in thalamic nuclei, both in health and disease.Acknowledgements
The authors would like to thank Stefano Tambalo for his support with anatomical data processing.
References
- Sherman, S.M. (2017). Functioning of circuits connecting thalamus and cortex. Comprehensive Physiology, 7(2), 713–739.
- Morel, A., Magnin, M., Jeanmonod, D. (1997). Multiarchitectonic and stereotactic atlas of the human thalamus. Journal of Comparative Neurology, 387(4), 588-630.
- Clascà, F., Rubio-Garrido, P., Jabaudon, D. (2012). Unveiling the diversity of thalamocortical neuronal subtypes. European Journal of Neuroscience, 35(10), 1524-1532.
- Sherman, S.M., & Guillery, R.W. (2006). Exploring the thalamus and its role in cortical function (2nd ed.). Cambridge, MA: MIT Press.
- Iglesias, J.E., Insausti, R., Lerma-Usabiaga, G., et al. (2018). A probabilistic atlas of the human thalamic nuclei combining ex vivo MRI and histology. NeuroImage, 183, 314-326.
- Jensen, J.H., Helpern, J.A., Ramani, A., Lu, H., Kaczynski, K. (2005). Diffusional kurtosis imaging: the quantification of non-gaussian water diffusion by means of magnetic resonance imaging. Magnetic Resonance in Medicine, 53(6), 1432-1440.
- Henriques, R.N., Correia, M.M., Marrale, M., et al. (2021). Diffusional kurtosis imaging in the diffusion imaging in Pyhton project. Frontiers in Human Neuroscience, 15, 675433.
- Veraart, J., Novikov, D.S., Christiaens, D., Ades-Aron, B., Sijbers, J., Fieremans, E. (2010). Denoising of diffusion MRI using random matrix theory. NeuroImage, 142, 394-406.
- Kellner, E., Dhital, B., Kiselev, V.G., Reisert, M. (2016). Gibbs‐ringing artifact removal based on local subvoxel‐shifts. Magnetic Resonance in Medicine, 76(5), 1574-1581.
- Andersson, J.L., & Sotiropoulos, S.N. (2016). An integrated approach to correction for off-resonance effects and subject movement in diffusion MR imaging. NeuroImage, 125, 1063-1078.
- R, v. 3.6.1, 2019-07-05, The R Foundation.
- Krauth, A., Blanc, R., Poveda, A., et al. (2010). A mean three-dimensional atlas of the human thalamus: Generation from multiple histological data. NeuroImage, 49, 2053–2062.
- Gao, Y., Schilling, K.G., Stepniewska, I., et al. (2018). Test of clustering thalamic nuclei based on various dMRI models in the squirrel monkey brain. Proceedings in SPIE, 10578.
- Beaulieu, C. (2002). The basis of anisotropic water diffusion in the nervous system - a technical review. NMR in Biomedicine, 15(7-8), 435-55.
- Friedrich, P., Fraenz, C., Schlüter, C., et al. (2020). The relationship between axon density, myelination, and fractional anisotropy in the human corpus callosum. Cerebral Cortex, 30, 2042:2056.
- Henriques, R.N., Jespersen, S.N., Shemesh, N. (2020). Correlation tensor magnetic resonance imaging. NeuroImage, 211, 116605.
- Steven, A.J., Zhuo, J., Melhem, E.R. (2014). Diffusion kurtosis imaging: an emerging technique for evaluating the microstructural environment of the brain. American Journal of Roentgenology, 202, W26-W33.
- Novello, L., Henriques, R.N., Ianuş, A., et al. (2021). In vivo Correlation Tensor MRI reveals microscopic kurtosis in the human brain on a clinical 3T scanner. bioRxiv, doi: https://doi.org/10.1101/2021.11.02.466950
- Borst, A., & Theunissen, F.E. (1999). Information theory and neural coding. Nature Neuroscience, 2, 947-957.
Figures