1711
Anisotropic transverse relaxation in the human brain white matter induced by restricted rotational diffusion
Yuxi Pang1
1Dept. of Radiology, University of Michigan, Ann Arbor, MI, United States
1Dept. of Radiology, University of Michigan, Ann Arbor, MI, United States
Synopsis
Orientation-dependent transverse ($$$R_2$$$ and $$$R_2^*$$$) relaxation phenomena have been documented in the human brain white matter, yet the underlying relaxation mechanisms still remain not well understood. This work is to propose an alternative relaxation pathway through restricted molecular rotational diffusion, in terms of a generalized magic angle effect (gMAE) model, to better characterize recently reported anisotropic $$$R_2$$$ of myelin water and intra- and extracellular water in vivo at 3T. The proposed gMAE model is intrinsically connected with anisotropic translational diffusion from DTI, elucidating not only previously reported $$$R_2^*$$$ anisotropy ex vivo but also its temperature-dependence at 7T.
INTRODUCTION
In highly ordered biological tissues, water proton MR transverse $$$T_2$$$ (i.e. 1/$$$R_2$$$) relaxation becomes orientation-dependent.1 In the past, several anisotropic $$$R_2$$$ and $$$R_2^*$$$ studies of the human brain white matter (WM) in vivo and ex vivo have been reported at $$$B_0$$$≥3T, and some relaxation mechanisms particularly including magnetic susceptibility and its anisotropy have been proposed.2-4 However, none of these relaxation mechanisms can fully explain the measured $$$R_2$$$ orientation-dependence at 3T and anisotropic $$$R_2^*$$$ temperature-dependence at 7T. Therefore, this work is to propose an alternative relaxation pathway to better characterize anisotropic transverse relaxation in WM.METHODS
(1) Theory: Given an image voxel comprising preferentially orientated myelin fibers, the bound water molecules are assumingly distributed in an axially symmetric system5, where an illustrative intramolecular dipole-dipole interaction vector <H-H> forms angle $$$\theta$$$ to $$$B_0$$$, and angle $$$\alpha$$$ to the symmetric axis ($$$\overrightarrow{n}$$$) that makes angle $$$\epsilon$$$ with $$$B_0$$$ (Figure 1A). Note, $$$\alpha$$$ and $$$\epsilon$$$ are stationary but $$$\varphi$$$ (azimuthal angle) and $$$\theta$$$ become time-dependent. When evaluated in this particular system, the orientation-dependence factor $$$\langle\left(3\cos^2\theta-1\right)^2\rangle$$$ of $$$R_2$$$ is called herein a generalized magic angle effect (gMAE) function. Because of the spherical law of cosines (i.e. $$$\cos\theta=\cos\alpha\cos\epsilon+\sin\alpha\sin\epsilon\cos\varphi$$$), the gMAE function can be expressed by either Eq. 1 or Eq. 2, after an ensemble averaging over $$$\varphi$$$ from 0 to 2π. $$f(α,ε)=(1/4)(3cos^2α-1)^2(3cos^2 ε-1)^2+(9/8)(sin^4αsin^4 ε+sin^22αsin^2 2ε) \; (1) $$ $$g(α,ε)=(1/512)(C_0(α)+C_1(α)cos2ε+C_2(α)cos4ε) \; (2) $$ The two equations appear markedly different; however, they provide the same results. Note, Eq. 2 was taken from Berendsen’s seminal work5, which is surprisingly identical to previously used models for characterizing $$$R_2$$$ and $$$R_2^*$$$ anisotropies.2-4 When $$$\alpha$$$=0°, $$$f(\alpha,\epsilon)$$$ returns to the standard MAE function. (2) Anisotropic $$$R_2$$$ in myelin water imaging at 3T: Anisotropic $$$R_2$$$ (black triangles) and its best fits (“Fit B”, dashed blue lines) from myelin water and intra- and extracellular (IE) water were extracted from a recent publication2, and reproduced in Figure 2A (myelin water) and 2B (IE water). (3) DTI of WM at 3T: An anisotropic translational diffusion ratio, i.e. $$$ADC_{\parallel}/ADC_{\perp}$$$=3.33±1.29 (mean ± SD), was retrieved from the literature6, indicating that water diffused, on average, along a direction deviated from $$$ADC_{\parallel}$$$ (or myelin fibers) direction with an angle $$$φ_1$$$=16.7±6.1°, as sketched in Figure 3B. (4) Anisotropic $$$R_2^*$$$ of the human brain specimens at 7T: The observed anisotropic $$$R_2^*$$$ data (Figure 4) were simulated (filled black circles) for two ROIs (green and magenta), based on the previous model $$$R_2^*(ε)= C_0+C_1sin(ε)+C_2sin(2ε-π⁄2)$$$ with $$$ε=2θ+φ_0$$$, and the best fitted model parameters.3 Additionally, temperature-dependent anisotropic $$$R_2^*$$$ were also restored4 and presented in Figure 5A. (5) Nonlinear least-squares fitting: The proposed $$$R_2$$$ model (Eq. 3) was fitted (“Fit A”) to the published anisotropic $$$R_2$$$ and $$$R_2^*$$$, with a phase shift ($$$φ_0$$$) to account for any directional differences between the axis ($$$\overrightarrow{n}$$$) and translational diffusion direction. $$R_2=R_2^i+R_2^a*f(α,ε-φ_0)\; (3)$$ As an independent variable ranging from 0° to 90°, $$$\epsilon$$$ was identified by orientations of myelin fiber tracts that were in turn determined by $$$ADC_{\parallel}$$$.6 A goodness of fit was indicated by the root-mean-square error (RMSE), and its statistical significance was assessed by an F-test, with significance indicated by P<.05. All data analysis was performed with in-house software written in IDL 8.5 (Harris Geospatial Solutions, Inc., Broomfield, CO, USA).RESULTS
Figures 1B-D show respectively a specific bound water distribution when $$$\alpha$$$=90°, a symmetric $$$f(α,ε)$$$ orientation dependence map, and 4 different $$$f(α,ε)$$$ profiles with $$$\alpha$$$=0° (red), 30° (green), 60° (blue) and 90° (black). Compared with the reported results in Figures 2A-B, the gMAE model (red solid lines) provided better fits for anisotropic $$$R_2$$$ in the human brain WM in vivo at 3T, as indicated by significantly reduced RMSEs for both myelin (2C, 0.352 vs. 0.767, P=.007) and IE water (2D, 0.015 vs. 0.031, P=.03). The fitting results show the comparable phase shifts $$$φ_0$$$ (15.1±1.1° vs. 16.9±1.1°) and angles $$$α$$$ (67.0±0.6° vs. 69.1±0.3°) but differing $$$R_2^i$$$ and $$$R_2^a$$$ within and between myelin and IE water. Interestingly, an average phase shift $$$φ_0$$$ from the gMAE model (Figure 3A) was approximately equal to $$$φ_1$$$ derived from DTI (Figure 3B), suggesting that anisotropic translational and rotational diffusions occur, on average, along the same direction in WM. Not only can the proposed gMAE model (solid lines) completely reproduce the simulated (black circles) anisotropic $$$R_2^*$$$ at 7T (Figure 4), but it also can account for, at least partially, the reported temperature-dependent changes of $$$R_2^*$$$ anisotropy (i.e. $$$∆R_2^*=R_{2\perp}^*-R_{2\parallel}^*$$$, Figure 5A) if $$$α$$$ decreases (to be more isotropic) from 75° (gold) to 65° (blue) when temperature increases as shown in Figure 5B.DISCUSSION
The introduced theoretical framework in this work is not new5, yet it possesses an enhanced explanatory power, relative to the previous models2-4, to account for $$$R_2$$$ and $$$R_2^*$$$ anisotropies in the human brain WM. The fact that the same orientation-dependent function (i.e. Eq. 2) can be derived from markedly different relaxation pathways is somewhat concerned, especially when inferring quantitative iron and myelin contents at higher $$$B_0$$$. Further research is warranted to separate the pertinent contributions to anisotropic transverse relaxation induced by different mechanisms particularly at higher $$$B_0$$$.CONCLUSION
The proposed generalized MAE model can better characterize $$$R_2$$$ and $$$R_2^*$$$ anisotropies in the human brain WM.Acknowledgements
No acknowledgement found.References
- Henkelman RM, Stanisz GJ, Kim JK, Bronskill MJ. Anisotropy of NMR properties of tissues. Magnetic resonance in medicine. 1994 Nov;32(5):592-601.
- Birkl C, Doucette J, Fan M, Hernandez-Torres E, Rauscher A. Myelin water imaging depends on white matter fiber orientation in the human brain. MRM 2020; doi: 10.1002/mrm.28543.
- Lee J, van Gelderen P, Kuo L-W, Merkle H, Silva AC, Duyn JH. T2*-based fiber orientation mapping. Neuroimage 2011;57(1):225-234.
- Oh S-H, Kim Y-B, Cho Z-H, Lee J. Origin of B0 orientation dependent R2*(= 1/T2*) in white matter. Neuroimage 2013;73:71-79.
- Berendsen HJC. Nuclear magnetic resonance study of collagen hydration. J Chem Phys 1962;36(12):3297-3305.
- Mädler B, Drabycz SA, Kolind SH, Whittall KP, MacKay AL. Is diffusion anisotropy an accurate monitor of myelination?: Correlation of multicomponent T2 relaxation and diffusion tensor anisotropy in human brain. Magnetic resonance imaging 2008;26(7):874-888.
Figures
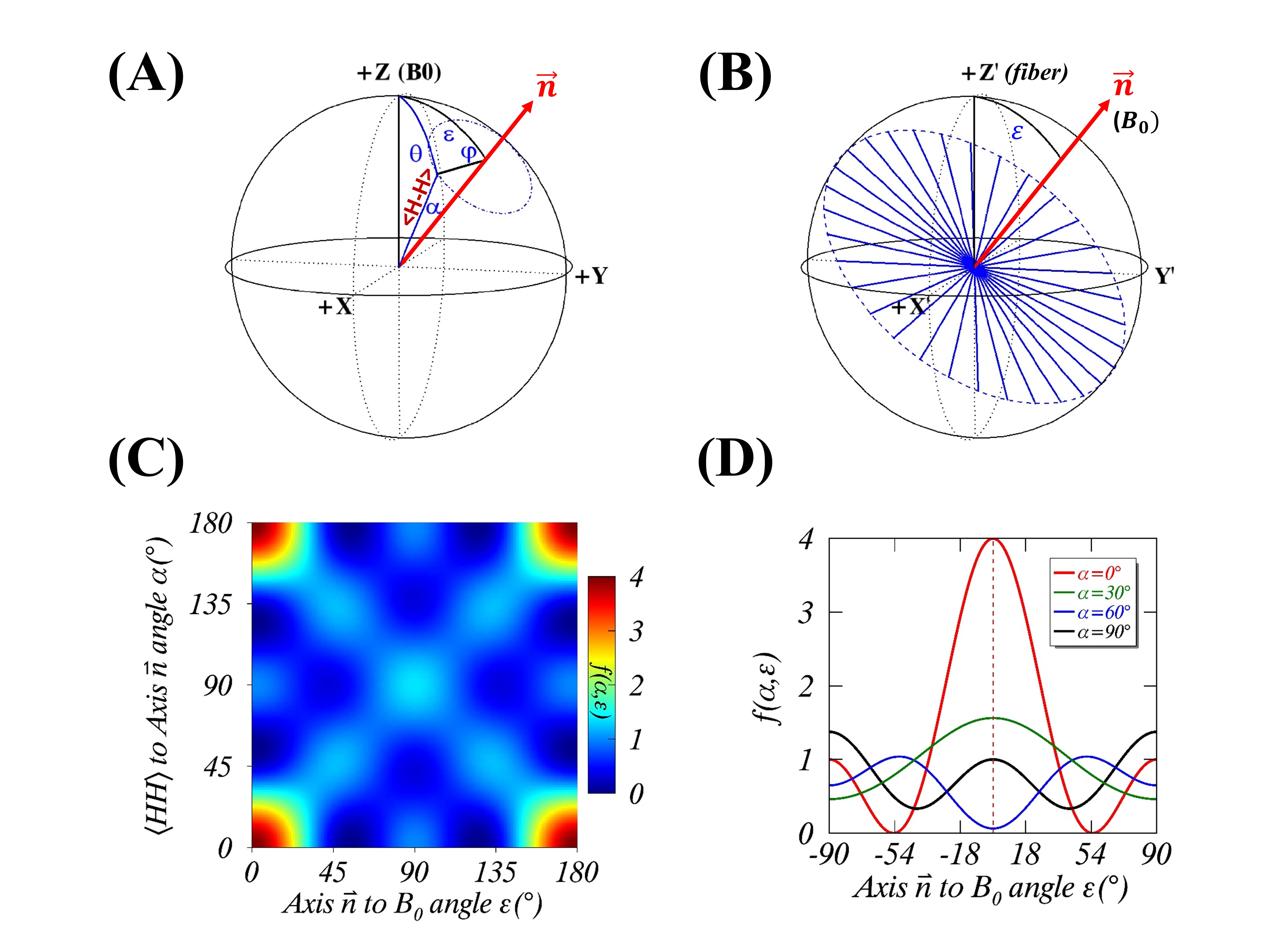
FIGURE 1. An axially symmetric rotational model for $$$R_2$$$ anisotropy in the magnet frame (A) with a specific distribution ($$$α$$$=90°) of water proton internuclear dipolar
interaction vectors <H-H> shown in the axonal fiber frame (B). A symmetric 2D map of
orientation-dependence $$$f(α,ε)$$$ is shown in
Figure 1C, and four representative $$$f(α,ε)$$$ profiles are
plotted in Figure 1D, for $$$α$$$=0° (red), 30° (green), 60° (blue) and 90° (black).
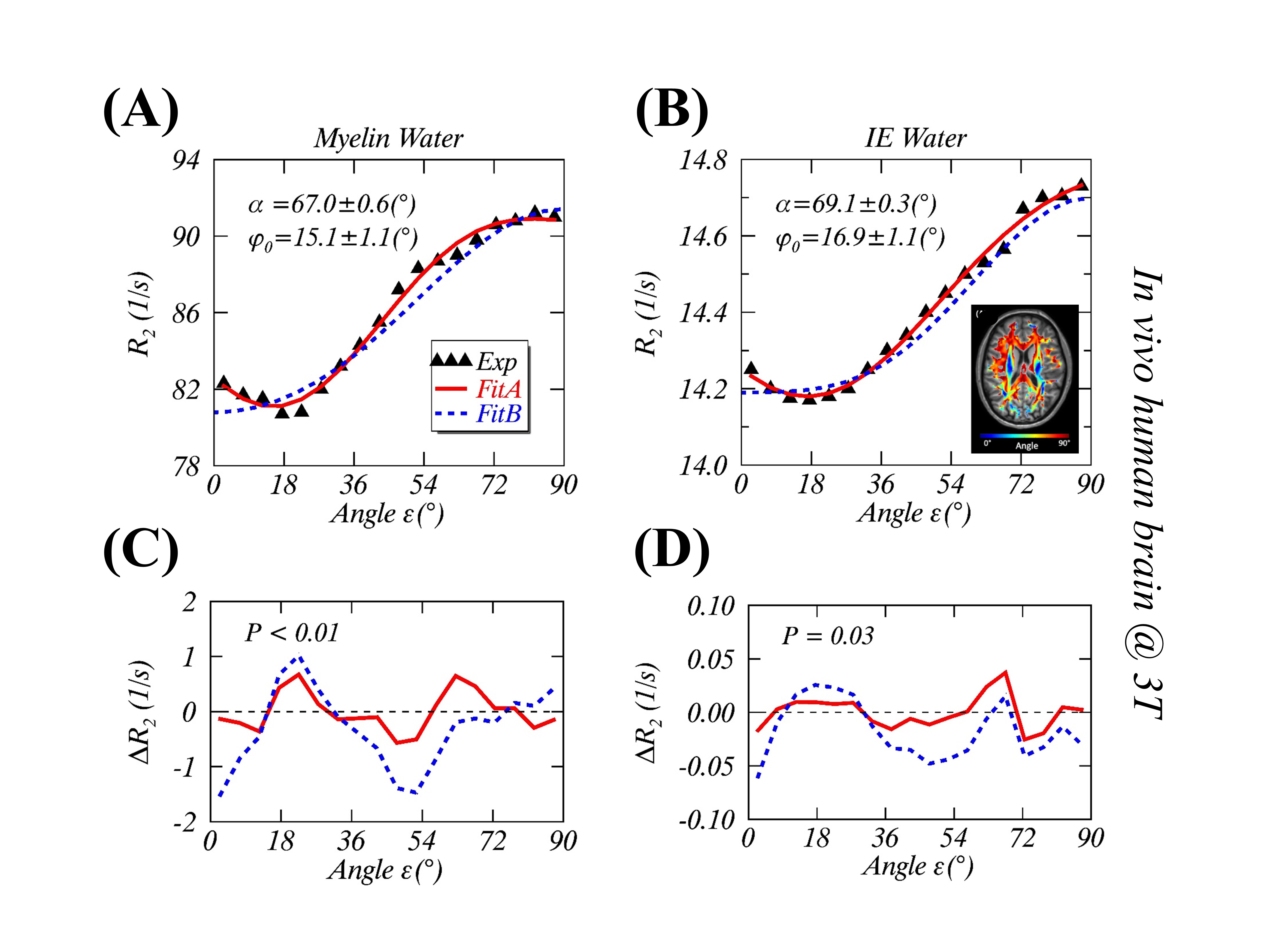
FIGURE 2. Measurements and modeling of anisotropic $$$R_2$$$ in the human
brain white matter in vivo at 3T. The measured
(black triangle) and the fitted anisotropic myelin water $$$R_2$$$ (2A) and
intra- and extracellular (IE) water $$$R_2$$$ (2B) using
the gMAE model with a phase offset $$$φ_0$$$ (Fit A, solid red lines), and the previous model (Fit B, dashed blue lines) without $$$φ_0$$$ from the
original publication [REF. 2]. The fitting residuals, defined as $$$ΔR_2$$$=Fitted-Measured, are respectively shown for myelin water (2C) and IE water (2D).
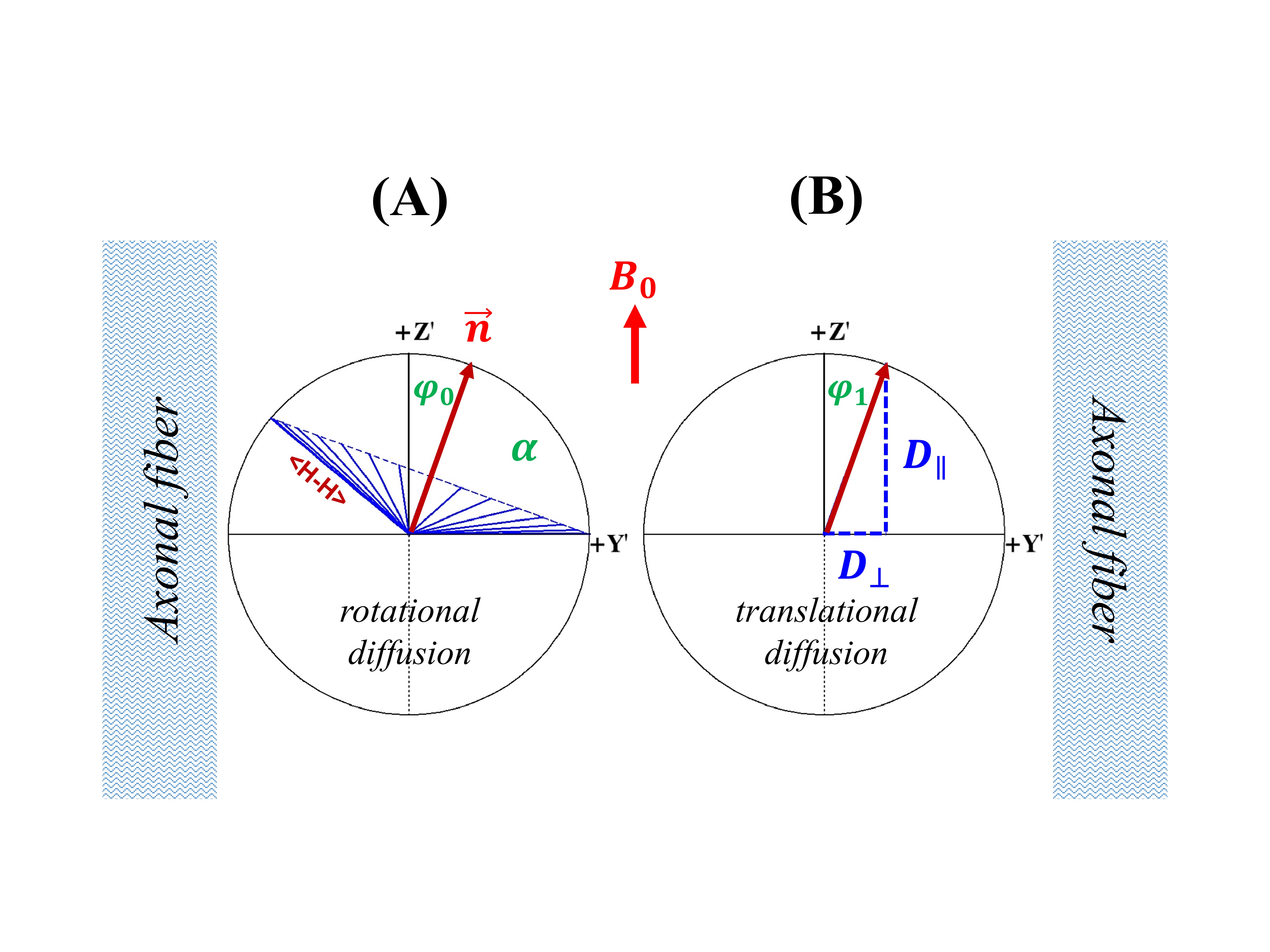
FIGURE 3. A schematic of water molecular restricted rotational diffusion (3A) and anisotropic translational diffusion (3B) in the human brain white matter.
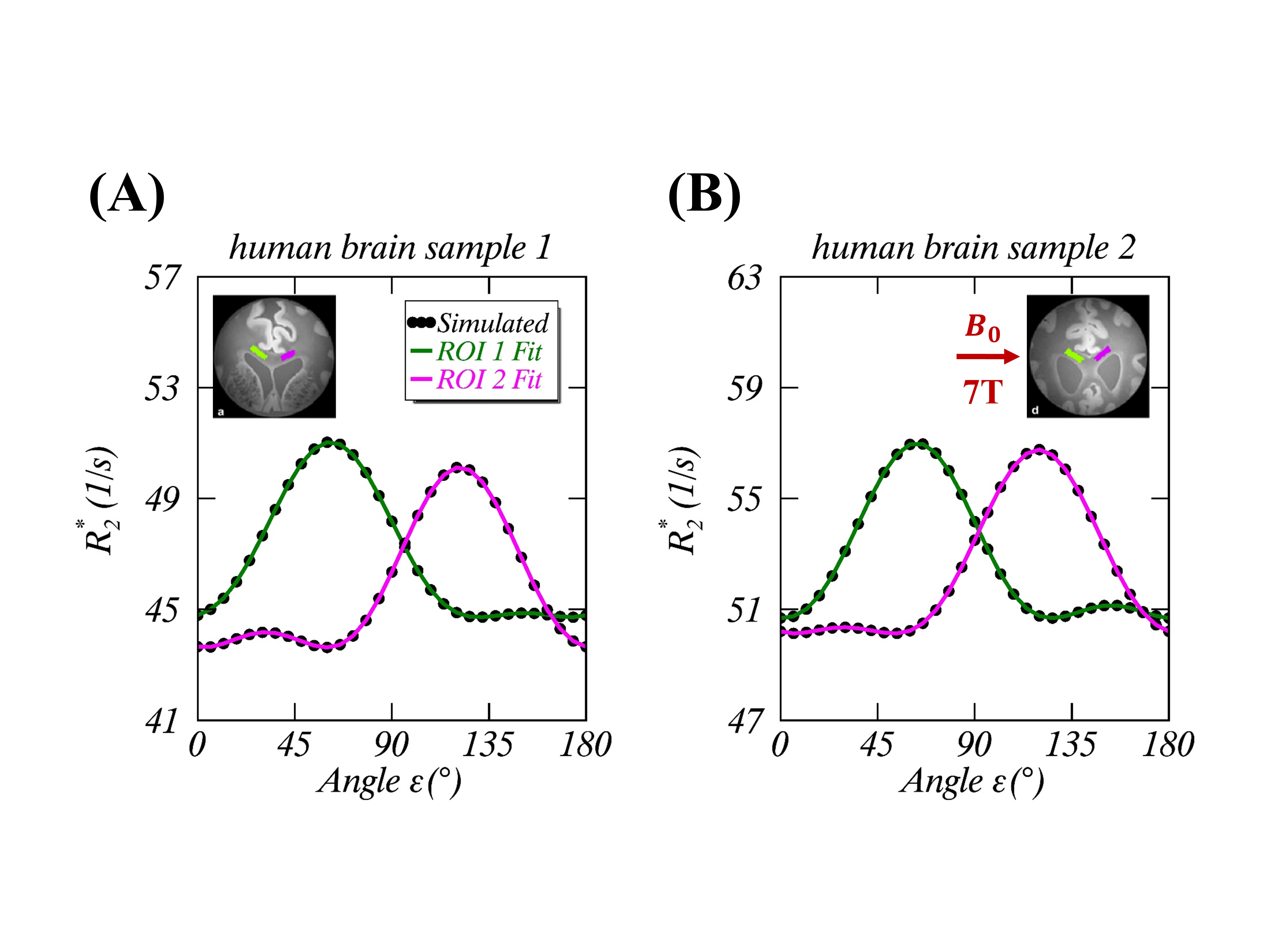
FIGURE 4. Simulated (black circles) and fitted (green lines for ROI1 and
magenta lines for ROI2) orientation-dependent $$$R_2^*$$$ at 7T from a
previous study [REF. 3] on two human brain specimens (sample 1 in Figure
4A and sample 2 in Figure 4B). Note, the simulations were carried out using a
previously published susceptibility anisotropy model along with the reported
fitted model parameters. The fitted lines shown in the graphs were obtained
with the gMAE model.
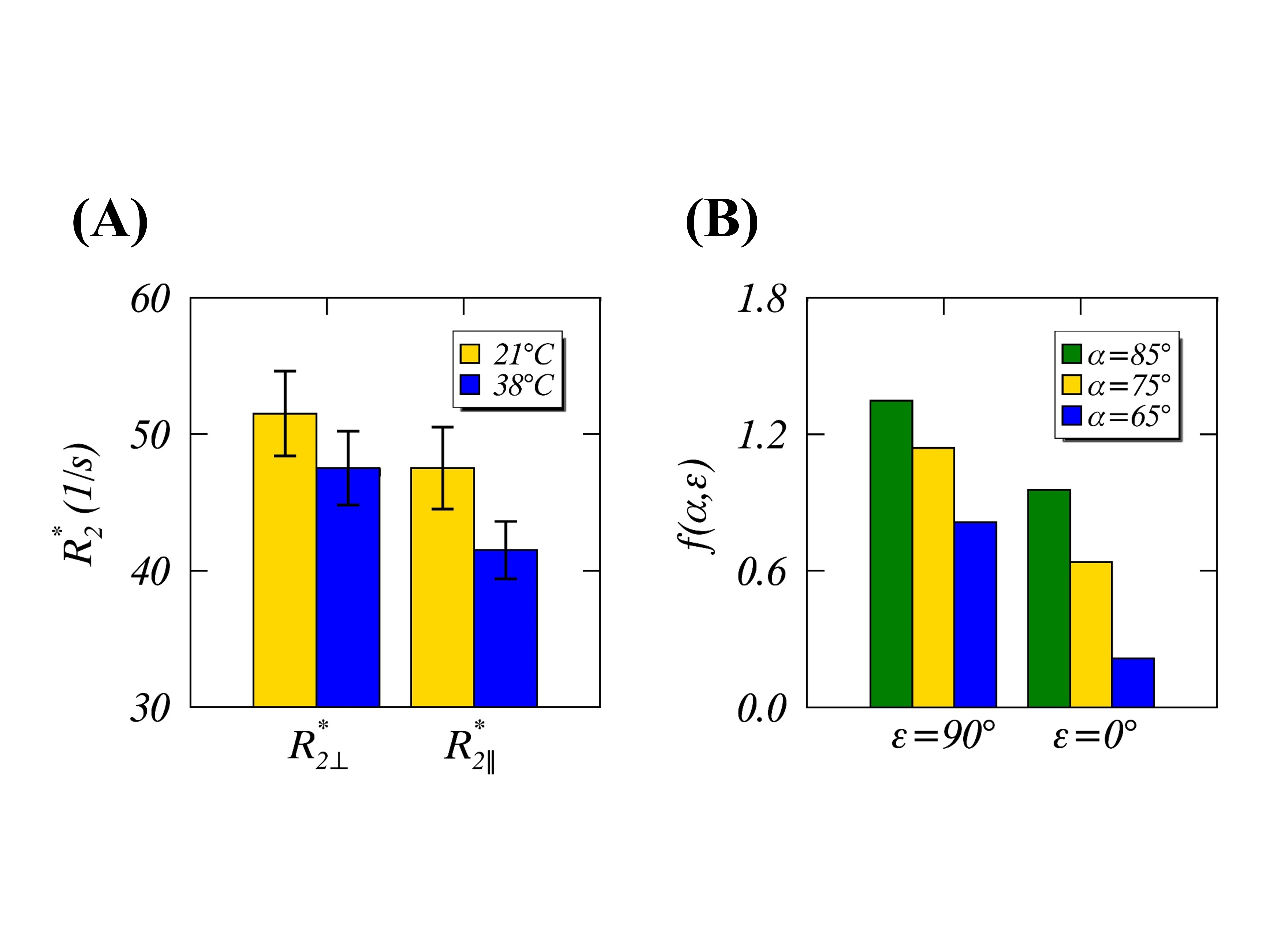
FIGURE 5. Reported temperature-dependent (21°C
in gold vs. 38°C in blue)
and orientation-dependent ($$$R_{2\perp}^*$$$ and $$$R_{2\parallel}^*$$$ when myelin
fibers perpendicular to and parallel to $$$B_0$$$, respectively) $$$R_2^*$$$ anisotropies
in the human brain specimens at 7T [REF. 4] (Figure 5A). Theoretical $$$f(α,ε)$$$ orientation-dependences by assuming myelin fibers perpendicular
to ($$$ε$$$=90°) and parallel ($$$ε$$$=0°) to $$$B_0$$$, when $$$α$$$=65° (blue),75° (gold), and 85° (green).