0676
Hyperpolarized [1-13C]pyruvate detects brain glucose metabolism and sex-specific vulnerability in glucose transporter deficient mice1Department of Physical Therapy and Rehabilitation Science, University of California San Francisco, San Francisco, CA, United States, 2Department of Radiology and Biomedical Sciences, University of California San Francisco, San Francisco, CA, United States, 3Gladstone Institute of Neurological Disease, San Francisco, CA, United States, 4Vanderbilt Center for Immunobiology, Department of Pathology, Microbiology, and Immunology, Vanderbilt University Medical Center, Nashville, TN, United States
Synopsis
We used hyperpolarized 13C magnetic resonance spectroscopic imaging (HP 13C MRSI), fluorine-18 fluorodeoxyglucose (18F-FDG) positron emission tomography (PET) imaging and T2-weighted MRI to detect brain glucose metabolism in mice harboring deletion of the glucose transporter 3 (GLUT3) in CA1 hippocampal neurons. GLUT3 deletion induced memory impairment in males and females, highlighting the importance of glucose uptake by neurons. HP [1-13C]lactate-to-pyruvate ratios and brain volumes were decreased in female GLUT3 deficient mice, but not in males, indicating sex-specific vulnerability. No changes were detected using 18F-FDG PET imaging, highlighting the potential of HP [1-13C]pyruvate to detect downstream alterations in brain glucose metabolism.
Introduction
The brain requires large amount of glucose, but it remains controversial whether neurons import it and further metabolize it through glycolysis, or if this role is essentially ensured by glial cells1,2,3. As disruption of glucose metabolism is an important feature of neurodegenerative disorders, it is of utmost importance to develop and validate noninvasive biomarkers to improve our knowledge of brain energetics in the healthy and diseased brain. In this context, animal models presenting deletion of key transporters and enzymes can provide mechanistic insights and well-controlled models to evaluate the potential of noninvasive methods to monitor metabolic changes in vivo. Positron emission tomography (PET) imaging using fluorine-18 fluorodeoxyglucose (18F-FDG) is well established to image glucose uptake, however one main limitation lies in its inability to inform on subsequent glycolytic steps and metabolic fluxes. Hyperpolarized 13C magnetic resonance spectroscopic imaging (HP 13C MRSI) has demonstrated great potential to measure metabolic fluxes in real-time in neurological disorders4-9. Specifically, HP [1-13C]pyruvate detects [1-13C]lactate production in the brain, thus informing on the metabolic fate of pyruvate, the end of product of glycolysis. Here, we used mice presenting a deletion of the glucose transporter 3 (GLUT3) in neurons of the CA1 hippocampal region, a key structure involved in memory formation. We investigated whether GLUT3 deletion leads to behavioral deficits and whether PET and MR imaging can detect in vivo GLUT3 alterations.Methods
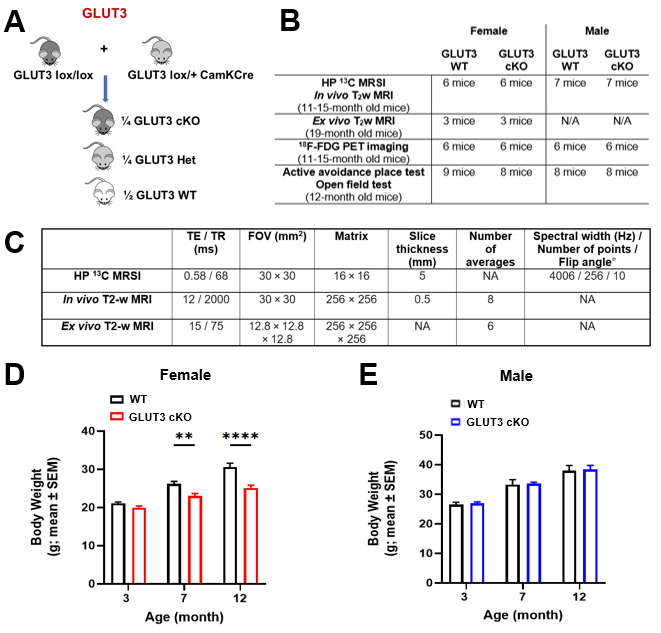
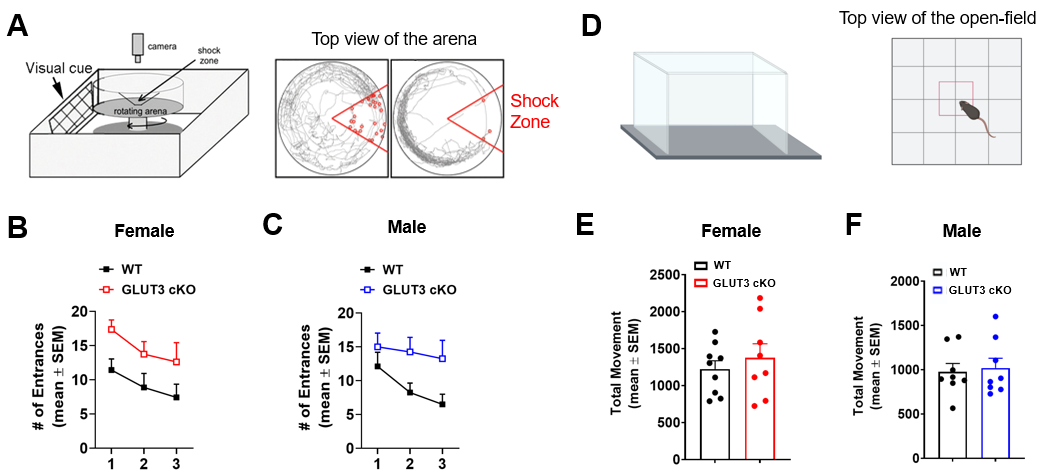
Animals: Floxed GLUT3 mice were bred with CamKII alpha (CamKCre) mice, which express Cre recombinase in nearly all CA1 neurons (Figure 1A), to obtain GLUT3 cKO and littermate GLUT3 WT controls. Mice underwent MRI, PET imaging and behavioral testing as shown in Figure 1B.Behavioral analyses: The active place avoidance test was used to assess hippocampal-dependent spatial learning. Using visual cues, mice learn to avoid the shock zone of a rotating arena. The number of entrances into the shock zone was calculated. The open-field test was used to measure locomotor and exploratory behavior, and the total movement over a 15-minute period was calculated.
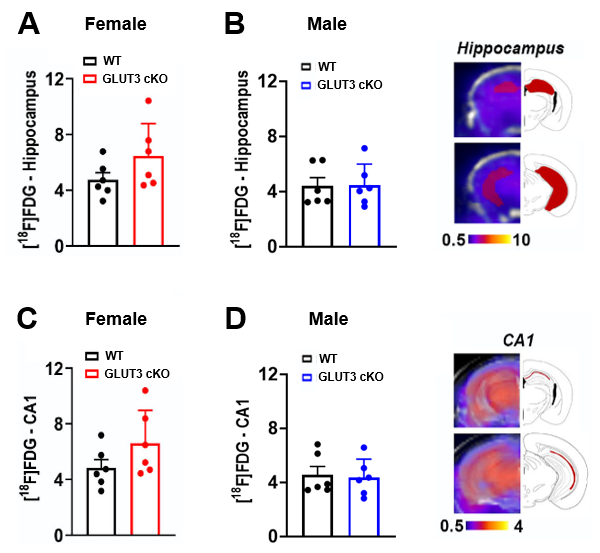
PET/computerized tomography (CT) acquisitions and analyses: PET/CT was acquired 55 minutes after 18F-FDG intravenous injection (71±4.5μCi). We co-registered PET and T2w MR images using VivoQuant software to delineate the hippocampus and CA1 region based on MR contrast and calculated the corresponding mean percent-injected dose per grams (%ID/g) values.
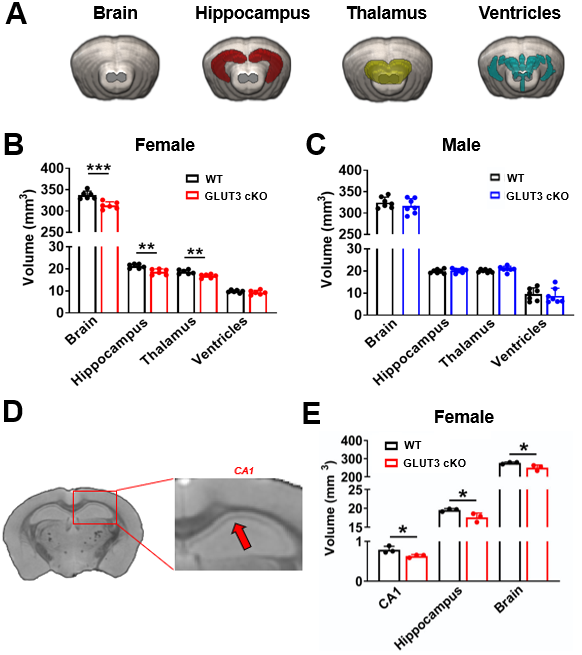
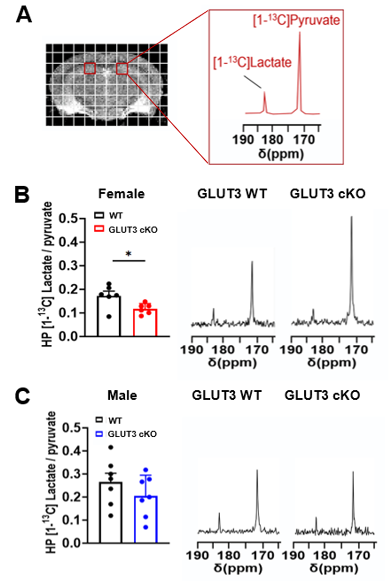
MR acquisitions and analyses: T2-weighted MRI and HP 2D 13C CSI were acquired on a 14.1T MR scanner using the parameters shown in Figure 1C. For 13C MRS, 24μl [1-13C] pyruvate was polarized for ~1h in a Hypersense polarizer, dissolved in 4.5mL buffer (80mM NaOH in PBS), and data were acquired 18 seconds after intravenous injection. HP 13C MRSI data were analyzed using the SIVIC software and MATLAB. K-space dimensions were zero-filled by two. The area-under-the-curve of HP [1-13C]pyruvate and [1-13C]lactate Lorentzian fits were measured and [1-13C]lactate/pyruvate ratios were calculated. For analyses of T2w MRI data, brain, thalamus, ventricles, hippocampus and CA1 regions were delineated and the corresponding volumes were calculated.
Statistical analyses: Two-way ANOVA or unpaired Student’s t-test were used to compare GLUT3WT and GLUT3cKO groups using GraphPad Prism software. *p<0.05, **p<0.01, ***p<0.001, ****p<0.001.
Results
Body weights were significantly decreased in female GLUT3cKO starting at 7-month-old mice (p=0.0048), while no changes were observed in males (Figure 1D-E). To determine if GLUT3 is required for normal hippocampal function, we examined how GLUT3cKO impacts spatial learning and memory assessed by active place avoidance. Higher number of entrances into the aversive shock zone reflected memory deficits in female and male GLUT3cKO (Figure 2A-C p=0.0451 and p=0.0492, respectively). No changes in total movement measured in the open field indicated no deficits in motor or sensory function (Figure 2D-F). Next, in vivo T2w anatomical MRI revealed smaller brain, hippocampus and thalamus in female GLUT3cKO (Figure 3A-B, p=0.0009, p=0.0033, p=0.0045, respectively), while ventricle size remained similar. No changes in brain regions were detected in males (Figure 3C). Ex vivo T2w MRI confirmed these results and additionally revealed smaller CA1 volume (Figure 3D-E, p=0.0458). Following injection of HP [1-13C]pyruvate, we observed production of [1-13C]lactate in the brain (Figure 4A). In the region containing hippocampus and CA1, we observed lower HP [1-13C]lactate-to-pyruvate ratios in female GLUT3cKO (p=0.0282), whereas this difference was not seen in males (Figure 4B-C). Interestingly, [18F]FDG PET signal from the hippocampus and CA1 were not significantly different between female or male GLUT3cKO and WT (Figure 5A-D).Discussion
We showed that GLUT3 deletion in CA1 hippocampal neurons led to memory impairment in male and female mice. Interestingly, anatomical and metabolic MRI, but not [18F]FDG PET imaging, detected changes linked to GLUT3 deletion in female mice only, highlighting sex-specific alterations. Furthermore, decreases in body weights and brain regions volumes in females suggest that GLUT3 deletion can cause systemic changes, representing an interesting area for future investigation. Altogether, our findings show that neurons import glucose to maintain normal function. Moreover, HP 13C-MRSI supports that glucose is metabolized through glycolysis in vivo, although there are gender differences in the reliance on GLUT3. Importantly, these results highlight the potential of HP [1-13C]pyruvate as a novel method to detect downstream alterations in brain glucose metabolism that were otherwise undetected using [18F]FDG PET imaging.Acknowledgements
CG, HL, KN and MMC contributed equally to this work. CG, HL, LLP, KN and MMC were supported by NIH 1RF1AG064170-01 to KN and MMC. This work was also supported by a grant from the California Department of Public Health (KN) and fellowships from the Larry L. Hillblom Foundation (LS) and the Alzheimer’s Association (HL and LLP).References
- 1. Patel, A. B. et al. Direct evidence for activity-dependent glucose phosphorylation in neurons with implications for the astrocyte-to-neuron lactate shuttle. Proc Natl Acad Sci U S A 111, 5385-5390, doi:10.1073/pnas.1403576111 (2014).
- 2. Lundgaard, I. et al. Direct neuronal glucose uptake heralds activity-dependent increases in cerebral metabolism. Nat Commun 6, 6807, doi:10.1038/ncomms7807 (2015).
- 3. Bélanger, M., Allaman, I. & Magistretti, P. J. Brain energy metabolism: focus on astrocyte-neuron metabolic cooperation. Cell Metab. 14, 724-738, doi:10.1016/j.cmet.2011.08.016 (2011).
- 4. Guglielmetti, C. et al. Hyperpolarized (13)C MR metabolic imaging can detect neuroinflammation in vivo in a multiple sclerosis murine model. Proc Natl Acad Sci U S A 114, E6982-E6991, doi:10.1073/pnas.1613345114 (2017).
- 5. Le Page, L. M., Guglielmetti, C., Taglang, C. & Chaumeil, M. M. Imaging Brain Metabolism Using Hyperpolarized (13)C Magnetic Resonance Spectroscopy. Trends Neurosci 43, 343-354, doi:10.1016/j.tins.2020.03.006 (2020).
- 6. Guglielmetti, C. et al. In vivo metabolic imaging of Traumatic Brain Injury. Sci Rep 7, 17525, doi:10.1038/s41598-017-17758-4 (2017).
- 7. DeVience, S. J. et al. Metabolic imaging of energy metabolism in traumatic brain injury using hyperpolarized [1-(13)C]pyruvate. Sci Rep 7, 1907, doi:10.1038/s41598-017-01736-x (2017).
- 8. Choi, Y. S. et al. Hyperpolarized [1-13C] pyruvate MR spectroscopy detect altered glycolysis in the brain of a cognitively impaired mouse model fed high-fat diet. Mol Brain 11, 74, doi:10.1186/s13041-018-0415-2 (2018).
- 9. Choi, Y. S. et al. Hyperpolarized [1-13C]lactate flux increased in the hippocampal region in diabetic mice. Mol Brain 12, 88, doi:10.1186/s13041-019-0505-9 (2019).
Figures