0576
Identifying functional correlations between lateral hypothalamus and cingulate cortex underlying brain state-dependent pupil dynamics1High-field Magnetic Resonance, Max Planck Institute for Biological Cybernetics, Tübingen, Germany, 2Graduate Training Centre of Neuroscience, Tübingen, Germany, 3Institute of Neuroscience and Medicine (INM-4), Forschungszentrum Jülich, Jülich, Germany, 4Athinoula A. Martinos Center, Massachusetts General Hospital and Harvard Medical School, Boston, MA, United States
Synopsis
The lateral hypothalamus (LH) is known to mediate different vigilance states and modulate pupil dilation through various neural populations. However, measuring subcortical neuronal activity non-invasively while assessing the brain state has remained challenging. Recently, it has been shown that the coupling between fMRI brain signals and pupil size fluctuations depends on the underlying brain state. In this work, we suggest that the synchronization of LH fMRI signals with pupil fluctuations may indicate modulation of the vigilance level of the brain.
Introduction
Animals under anesthesia show altered brain states mimicking different stages of sleep. The lateral hypothalamus (LH) plays an important role in alternating brain states, in particular, mediating vigilance levels during rapid eye movement (REM)1 or non-REM (NREM)2 sleep and wakefulness3. Meanwhile, pupil dynamics have been often used as an arousal index to tract brain state changes in both anesthetized4 or awake animals5 and humans6. The noradrenergic projection circuit from the locus coeruleus (LC) to the anterior cingulate cortex (ACC) has been mostly investigated to interpret the brain state-dependent pupil dynamics7,8,9. Lately, we have detected that the neuronal oscillation of ACC shows both positive and negative correlation to pupil dynamics in anesthetized rats, which cannot be explained solely by the noradrenergic neuromodulation. The subcortical regulation of the opposite correlation features of ACC remains unknown. The LH has been reported to mediate both brain state changes10 and autonomic activity11, both of which could contribute to regulating pupil dynamics. We hypothesize that the LH interacts with the ACC to modulate the brain state-dependent pupil dynamics, presenting different correlation patterns. Here, we performed electrophysiological recordings in both LH and ACC and simultaneous fMRI and fiber photometry-based neuronal Ca2+ in the ACC to elucidate the neuronal basis of the opposite correlations to pupil dynamics in varied brain statesMethods
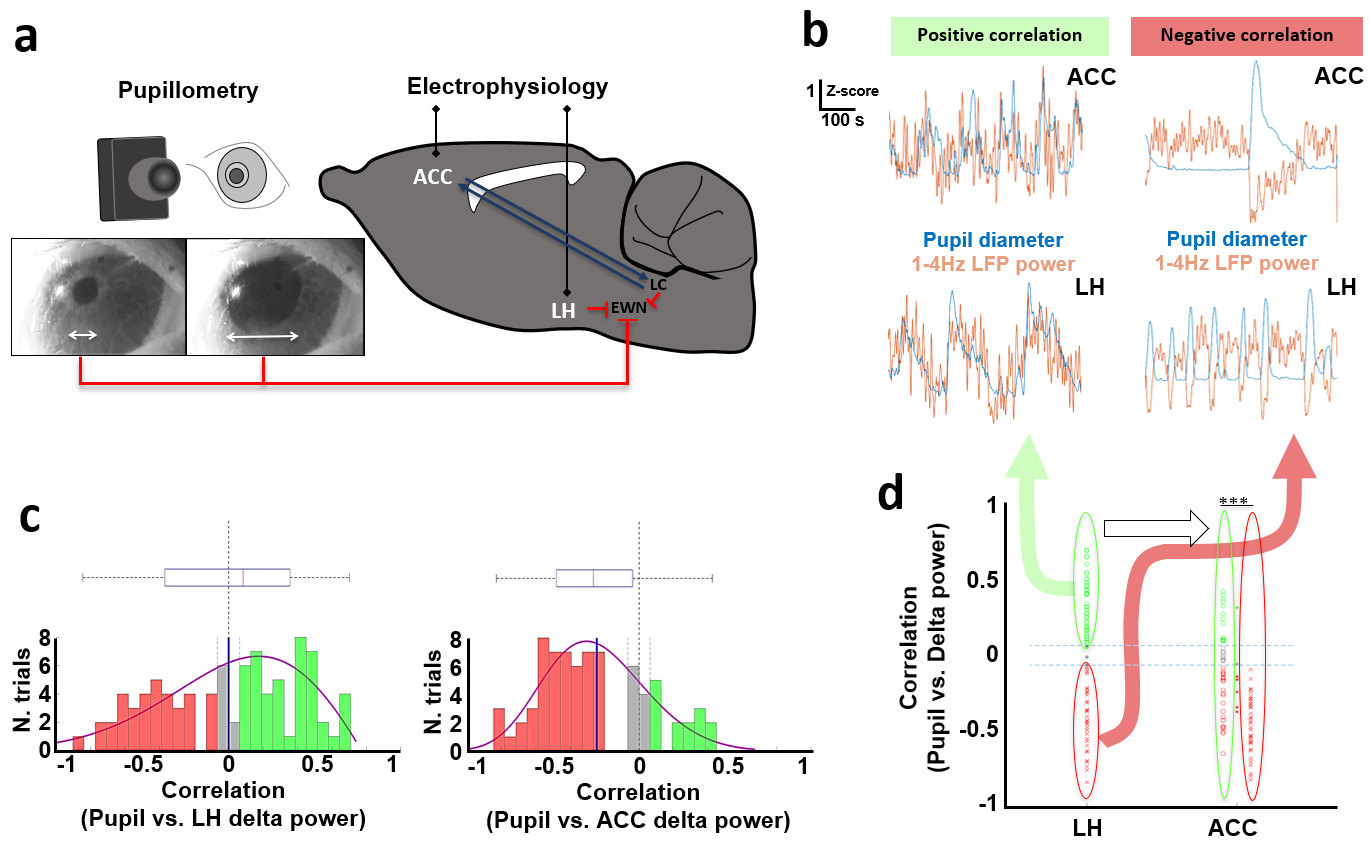
LFPs recording and pupil size detectionResting state electrophysiology signals were recorded from the LH and ACC simultaneously with pupil size detection in 15-min trials from nine rats anesthetized with alpha-chloralose (n= 89) (Fig 1a).
fMRI with concurrent pupil size detection and optical fiber calcium signal recording
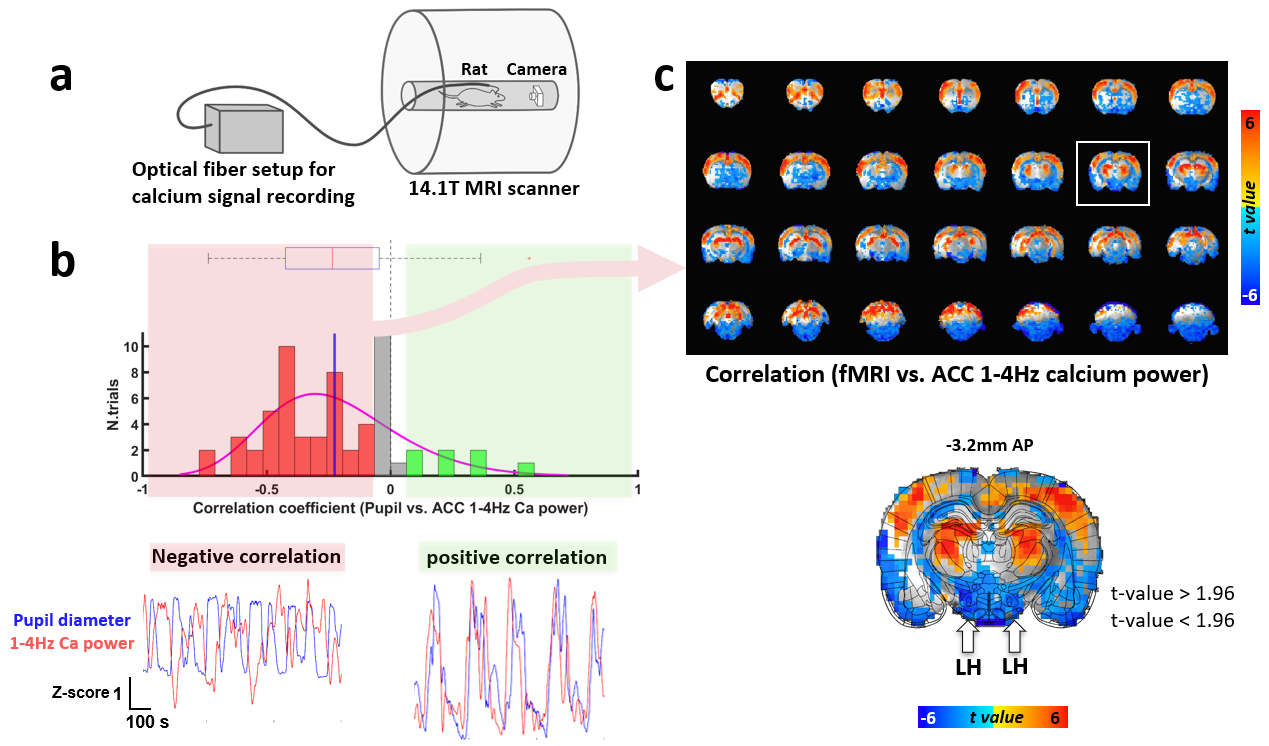
Resting state whole-brain fMRI and optical fiber calcium signal (GCaMP) recording from the ACC and pupil size detection were acquired in 15-min trials simultaneously from nine rats anesthetized with alpha-chloralose (n= 61) with a 14.1 T/26 cm magnet (Magnex), as previously described4 (Fig 3a).
Results
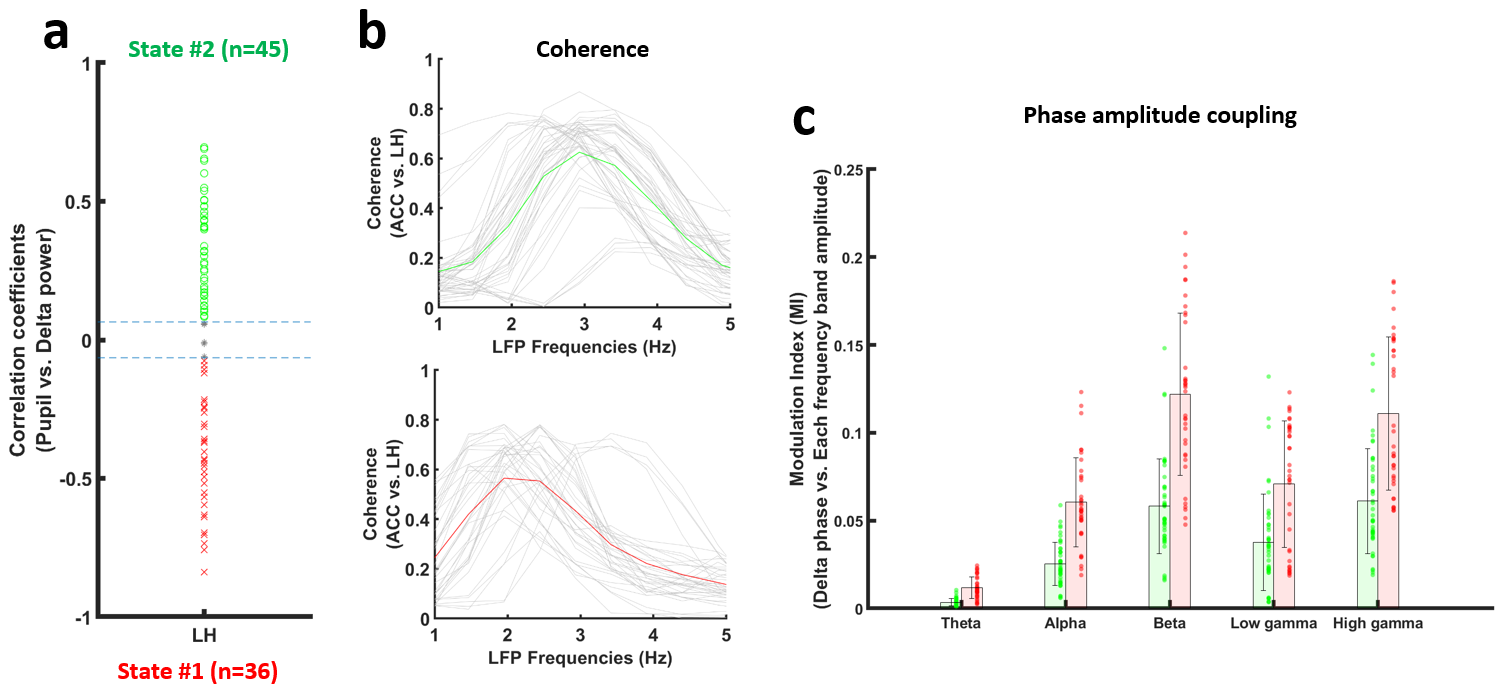
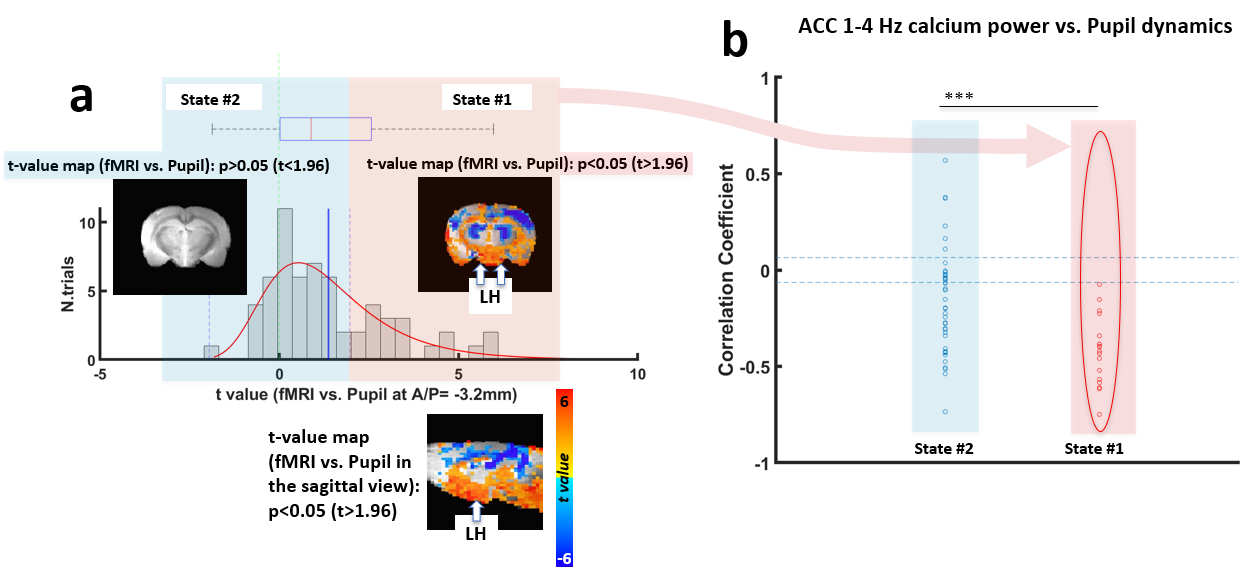
Electrophysiological recordings demonstrated that LFP delta (1-4Hz) power in both of the LH and ACC exhibited a positive or negative correlation with pupil size fluctuation (Fig 1b and 1c). When pupil dynamics and LH-delta power were anti-correlated (state #1), the correlation between pupil dynamics and ACC-delta power was always negative (Fig 1d). Under this state, the LH and ACC exhibited their highest coherence at 2Hz (Fig 2b). In contrast, when LH-delta power was positively correlated with the pupil dynamics (state #2), the highest coherence between the LH and ACC occurred at 3Hz (Fig 2b). In addition, phase-amplitude coupling at the ACC between the phase of delta and the amplitude of all other LFP frequency bands was observed to be higher in state #1 (Fig 2c). Thus, LH delta power presents a robust indicator to differentiate brain state-dependent correlation features to pupil dynamics. Similar to the electrophysiological signals in the ACC, 1-4Hz calcium power fluctuations in the ACC also showed positive or negative correlations with the pupil dynamics (Fig 3b). Interestingly, during the negative correlation state, ACC 1-4Hz calcium power was also negatively correlated with the fMRI signal in the LH (Fig 3c). Previous study also suggested a potential negative correlation of BOLD fMRI with delta power fluctuation in the subcortical region, e.g. striatum12. It is possible that the BOLD fMRI signal is negatively correlated with the delta power fluctuation in LH too. To examine whether the fMRI signal in the LH could differentiate brain-state dependent pupil dynamics similar to the electrophysiological signals, we correlated the pupil dynamics with the whole-brain fMRI, showing both positive and negative correlation in LH region (Fig 4a). Importantly, when categorizing trials showing positive correlation between pupil dynamics and LH fMRI signal (Fig 4a), we detected robust negative correlations between ACC 1-4Hz calcium power and pupil dynamics (Fig 4b), which is similar to the electrophysiological results (Fig 1d). This result indicates that the LH fMRI signal can also be used to differentiate brain states, presenting opposite correlation with pupil dynamics to delta power in the LH.Discussion and Conclusion
We analyzed the electrophysiological signal (in particular, the delta power) and fMRI signal in the LH together with pupil dynamics, showing a strong indicator of different brain states represented by positive and negative correlations of ACC activity and pupil dynamics. Interestingly, when LH delta power is negatively correlated with pupil dynamics, a 2Hz dominant brain state is observed, which is consistent with the NERM condition13. In contrast, when the LH delta power is positively correlated with pupil dynamics, a 3Hz dominant state is more correlated with a REM state during sleep14. Meanwhile, besides the electrical signals recorded from the LH, we could use the fMRI signal from the LH to determine the different brain state, but showing the opposite correlation pattern with electrophysiology.Acknowledgements
This work was supported by Max Planck Society and the Graduate Training Center of Neuroscience, International Max Planck Research School.References
1. Hassani OK, Lee MG, Jones BE. Melanin-concentrating hormone neurons discharge in a reciprocal manner to orexin neurons across the sleep-wake cycle. Proc Natl Acad Sci U S A. 2009;106(7):2418-22.
2. Blanco-Centurion C, Liu M, Konadhode RP, Zhang X, Pelluru D, van den Pol AN, Shiromani PJ. Optogenetic activation of melanin-concentrating hormone neurons increases non-rapid eye movement and rapid eye movement sleep during the night in rats. Eur J Neurosci. 2016;44(10):2846-2857.
3. Adamantidis AR, Zhang F, Aravanis AM, Deisseroth K, de Lecea L. Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature. 2007;450(7168):420-4.
4. Pais-Roldán P, Takahashi K, Sobczak F, Chen Y, Zhao X, Zeng H, Jiang Y, Yu X. Indexing brain state-dependent pupil dynamics with simultaneous fMRI and optical fiber calcium recording. Proc Natl Acad Sci U S A. 2020;117(12):6875-6882.
5. McGinley MJ, David SV, McCormick DA. Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection. Neuron. 2015;87(1):179-92.
6. Bradshaw J. Pupil size as a measure of arousal during information processing. Nature. 1967;216(5114):515-6.
7. Joshi S, Li Y, Kalwani RM, Gold JI. Relationships between Pupil Diameter and Neuronal Activity in the Locus Coeruleus, Colliculi, and Cingulate Cortex. Neuron. 2016;89(1):221-34.
8. Liu Y, Rodenkirch C, Moskowitz N, Schriver B, Wang Q. Dynamic Lateralization of Pupil Dilation Evoked by Locus Coeruleus Activation Results from Sympathetic, Not Parasympathetic, Contributions. Cell Rep. 2017;20(13):3099-3112.
9. Koga K, Yamada A, Song Q, Li XH, Chen QY, Liu RH, Ge J, Zhan C, Furue H, Zhuo M, Chen T. Ascending noradrenergic excitation from the locus coeruleus to the anterior cingulate cortex. Mol Brain. 2020;13(1):49.
10. Lee SH, Dan Y. Neuromodulation of brain states. Neuron. 2012;76(1):209-222.
11. Szabadi E. Functional Organization of the Sympathetic Pathways Controlling the Pupil: Light-Inhibited and Light-Stimulated Pathways. Front Neurol. 2018;9:1069.
12. Jaime S, Gu H, Sadacca BF, Stein EA, Cavazos JE, Yang Y, Lu H. Delta Rhythm Orchestrates the Neural Activity Underlying the Resting State BOLD Signal via Phase-amplitude Coupling. Cereb Cortex. 2019;29(1):119-133.
13. Benoit O, Daurat A, Prado J. Slow (0.7-2 Hz) and fast (2-4 Hz) delta components are differently correlated to theta, alpha and beta frequency bands during NREM sleep. Clin Neurophysiol. 2000;111(12):2103-6.
14. Bernardi G, Betta M, Ricciardi E, Pietrini P, Tononi G, Siclari F. Regional Delta Waves In Human Rapid Eye Movement Sleep. J Neurosci. 2019;39(14):2686-2697.
Figures