0043
Brain metabolic impairment after mild repetitive traumatic brain injury can be measured by hyperpolarized [1-13C]pyruvate and [13C]urea
Caroline Guglielmetti1,2, Kai Qiao1,2, Brice Tiret1,2, Karen Krukowski1,3, Amber Nolan3,4, Susanna Rosi1,3,5,6, and Myriam M. Chaumeil1,2
1Department of Physical Therapy and Rehabilitation Science, University of California San Francisco, San Francisco, CA, United States, 2Department of Radiology and Biomedical Sciences, University of California San Francisco, San Francisco, CA, United States, 3Brain and Spinal Injury Center, University of California San Francisco, San Francisco, CA, United States, 4Department of Pathology, University of California San Francisco, San Francisco, CA, United States, 5Department of Neurological Surgery, University of California San Francisco, San Francisco, CA, United States, 6Weill institute for Neuroscience, University of California San Francisco, San Francisco, CA, United States
1Department of Physical Therapy and Rehabilitation Science, University of California San Francisco, San Francisco, CA, United States, 2Department of Radiology and Biomedical Sciences, University of California San Francisco, San Francisco, CA, United States, 3Brain and Spinal Injury Center, University of California San Francisco, San Francisco, CA, United States, 4Department of Pathology, University of California San Francisco, San Francisco, CA, United States, 5Department of Neurological Surgery, University of California San Francisco, San Francisco, CA, United States, 6Weill institute for Neuroscience, University of California San Francisco, San Francisco, CA, United States
Synopsis
We used hyperpolarized 13C magnetic resonance spectroscopic imaging (HP 13C MRSI), T1- and T2-MRI to detect brain alterations in a mouse model of mild repetitive traumatic brain injury (rTBI). T1/T2-MRI did not detect brain damages. HP 13C MRSI detected metabolic changes in cortical areas, with decreased HP lactate/pyruvate and pyruvate dehydrogenase activity in rTBI. Interestingly, HP pyruvate and HP urea increased in rTBI, suggesting vascular and/or blood brain barrier alterations. Altogether, we demonstrated that HP 13C MRSI has potential to detect long-lasting metabolic alterations following rTBI and holds great potential for improving diagnosis and monitoring of rTBI in clinical practice.
Introduction
Individuals subject to frequent concussions such as football players or boxers, victims of abusive head trauma, or active military are some of the population at risk for repetitive mild traumatic brain injury (rTBI). rTBI is a being steadily recognized to increase the risk of neurodegenerative diseases, particularly the development of chronic traumatic encephalopathy (CTE). Alterations in prefrontal cortex-dependent functions including risk-taking phenotypes, social behavior, working and episodic memory have been reported at 1-3 months after five mild hits in mice1,2. Furthermore, long lasting hippocampal-dependent learning impairments have been reported up to 6 months after three mild hits in mice3, denoting the long-term effects of rTBI. However, the mechanisms underlying the adverse effects of rTBI are largely unknown. In the majority of cases, structural damage following rTBI is not seen on in vivo T2-weighted MR imaging1, hampering proper diagnosis and appropriate clinical care. There is thus a need for more sensitive noninvasive techniques to detect rTBI-induced brain changes. Hyperpolarized 13C magnetic resonance spectroscopic imaging (HP 13C MRSI) can measure metabolic fluxes in real time and has been shown to detect metabolic changes in moderate TBI models4,5. Here, we investigated whether 13C MRSI of HP [1-13C]pyruvate and [13C]urea could detect metabolic impairment in vivo in a mild rTBI model. T2-w MRI and T1-mapping were also performed to assess, respectively, potential structural changes and/or variations in T1 relaxation times due to tissue microstructure alterations and oxidative stress6.Methods
Animals and mild rTBI induction: Eight weeks old male C57BL/6J mice were assigned to a Sham (n=10) or rTBI (n=9) group. rTBI animals were subjected to multiple mild head injuries using the Closed-Head Impact Model of Engineered Rotational Acceleration (CHIMERA) device1,7. rTBI animals received a cortical injury (impact centered on the dorsal convexities of the skull, targeting a 5 mm area) once per day for 5 days with a 24-hour interval in between impacts. Sham mice were exposed to the procedure without sustaining an impact.MR acquisitions: T2-w MRI, T1-maps and HP 2D 13C CSI were acquired 3.5 months after rTBI (5.5-month-old mice), on a 14.1T MR Agilent scanner (Figure 1A), using the parameters shown in Figure 1B. For HP 13C data, 24μl [1-13C]pyruvate and 60μl [13C]urea were co-polarized for ~1h in a Hypersense polarizer and dissolved in 4.5mL buffer (80mM NaOH in PBS). 2D dynamic CSI 13C data were acquired 16 seconds post-intravenous injection.
Data analysis: Brain regions were manually delineated on T1 maps and T2-w images for each mouse based on the Allen Adult Mouse Brain atlas using the Aedes package for MATLAB. For each region, the volume was calculated using T2w data, and the mean T1 relaxation time using T1 maps. HP 13C MRSI data were analyzed using the SIVIC software and custom-built programs written in MATLAB. The area-under-the-curve of HP [1-13C]pyruvate, [1-13C]lactate and [13C]urea were measured for cortical (2 voxels/mouse) and subcortical areas (2 voxels/mouse), normalized to noise, and [1-13C]lactate/pyruvate ratios were calculated.
Spectrophotometric assays: Pyruvate dehydrogenase (PDH) activity was measured for cortical and subcortical regions.
Statistical analyses: Results are reported as mean±std. Unpaired Student t-test were used to compare Sham and rTBI groups using GraphPad Prism. *p<0.05, **p<0.01, ***p<0.001.
Results
No significant differences in volumes of whole brain and cerebral subregions could be detected on T2-w MRI between Sham and rTBI (Figure 2A). Based on T1 maps, no significant differences in T1 relaxation values were found between Sham and rTBI for any regions (Figure 2B).In cortical voxels, the level of HP [1-13C]lactate was comparable between Sham and rTBI groups (Figure 3A-B). However, HP [1-13C]pyruvate and HP [1-13C]urea were both significantly increased by 116±26% (p=0.0101) and 117±25% (p=0.006), respectively, in the rTBI group (Figure 3C-D). The subsequent HP [1-13C]lactate/pyruvate ratio was significantly lower in rTBI (Figure 3E, 87±18%, (p=0.001)). In subcortical voxels, all three HP metabolites were increased (Figure 3F-I, lactate by 113±20%, (p=0.0087); pyruvate by 118±29%, (p=0.0149), urea by 125±25%, (p=0.0002)). However, the HP [1-13C]lactate/pyruvate ratio was not significantly different between Sham and rTBI (Figure 3J, p=0.4462). Heatmaps of HP metabolites clearly indicate overall brain increase of HP pyruvate and urea, and lower HP lactate/pyruvate ratio in cortical areas in rTBI (Figure 4).
Pyruvate dehydrogenase (PDH), the enzyme that converts pyruvate into acetyl-coA and controls its entry into the tricarboxylic acid cycle, was decreased in cortical areas (Figure 5A-B, prefrontal cortex (p=0.0044), and cortex (p=0.0375)) of rTBI, but no changes were detected in subcortical areas (Figure 5C-D, hippocampus and thalamus).
Discussion
Although T2- and T1-MRI failed to detect injury, we observed metabolic alterations in cortical areas characterized by decreased HP lactate/pyruvate ratios and decreased PDH activity in rTBI. Decreased PDH activity has been linked to microglial activation in multiple sclerosis and moderate TBI models, however resulting in increased lactate/pyruvate ratios4,8. Interestingly, we observed higher HP pyruvate and urea levels in rTBI, suggesting vascular changes, and/or changes in blood brain barrier permeability9-12. Future studies are underway to dissect the mechanisms driving the HP readouts.Altogether our results demonstrate that HP 13C MRSI can detect metabolic impairment in vivo in a rTBI model, and thus offer new possibilities for the detection of rTBI-induced damages in the clinical setting.
Acknowledgements
This work was supported by research grants: NIH R01NS102156 (MC), NIH NIA grants R01AG056770 (SR) and NIH R21NS096718 (SR, MC), NMSS research grant RG-1701-26630 (MC), Dana Foundation: The David Mahoney Neuroimaging program (MC) and the NIH Hyperpolarized MRI Technology Resource Center #P41EB013598. K.K. is supported by an NRSA post-doctoral fellowship from the NIA F32AG054126.References
- 1. Nolan, A. et al. Repeated Mild Head Injury Leads to Wide-Ranging Deficits in Higher-Order Cognitive Functions Associated with the Prefrontal Cortex. J Neurotrauma 35, 2425-2434, doi:10.1089/neu.2018.5731 (2018).
- 2. Krukowski, K. et al. Integrated Stress Response Inhibitor Reverses Sex-Dependent Behavioral and Cell-Specific Deficits after Mild Repetitive Head Trauma. J Neurotrauma 37, 1370-1380, doi:10.1089/neu.2019.6827 (2020).
- 3. Chen, H., Desai, A. & Kim, H. Y. Repetitive Closed-Head Impact Model of Engineered Rotational Acceleration Induces Long-Term Cognitive Impairments with Persistent Astrogliosis and Microgliosis in Mice. J Neurotrauma 34, 2291-2302, doi:10.1089/neu.2016.4870 (2017).
- 4. Guglielmetti, C. et al. In vivo metabolic imaging of Traumatic Brain Injury. Sci Rep 7, 17525, doi:10.1038/s41598-017-17758-4 (2017).
- 5. DeVience, S. J. et al. Metabolic imaging of energy metabolism in traumatic brain injury using hyperpolarized [1-(13)C]pyruvate. Sci Rep 7, 1907, doi:10.1038/s41598-017-01736-x (2017).
- 6. Tain, R. W., Scotti, A. M., Li, W., Zhou, X. J. & Cai, K. Imaging short-lived reactive oxygen species (ROS) with endogenous contrast MRI. J Magn Reson Imaging 47, 222-229, doi:10.1002/jmri.25763 (2018).
- 7. Namjoshi, D. R. et al. Merging pathology with biomechanics using CHIMERA (Closed-Head Impact Model of Engineered Rotational Acceleration): a novel, surgery-free model of traumatic brain injury. Mol Neurodegener 9, 55, doi:10.1186/1750-1326-9-55 (2014).
- 8. Guglielmetti, C. et al. Hyperpolarized (13)C MR metabolic imaging can detect neuroinflammation in vivo in a multiple sclerosis murine model. Proc Natl Acad Sci U S A 114, E6982-E6991, doi:10.1073/pnas.1613345114 (2017).
- 9. Tagge, C. A. et al. Concussion, microvascular injury, and early tauopathy in young athletes after impact head injury and an impact concussion mouse model. Brain 141, 422-458, doi:10.1093/brain/awx350 (2018).
- 10. Veksler, R. et al. Slow blood-to-brain transport underlies enduring barrier dysfunction in American football players. Brain 143, 1826-1842, doi:10.1093/brain/awaa140 (2020).
- 11. Bashir, A. et al. Increased severity of the CHIMERA model induces acute vascular injury, sub-acute deficits in memory recall, and chronic white matter gliosis. Exp Neurol 324, 113116, doi:10.1016/j.expneurol.2019.113116 (2020).
- 12. Abrahamson, E. E. & Ikonomovic, M. D. Brain injury-induced dysfunction of the blood brain barrier as a risk for dementia. Exp Neurol 328, 113257, doi:10.1016/j.expneurol.2020.113257 (2020).
Figures
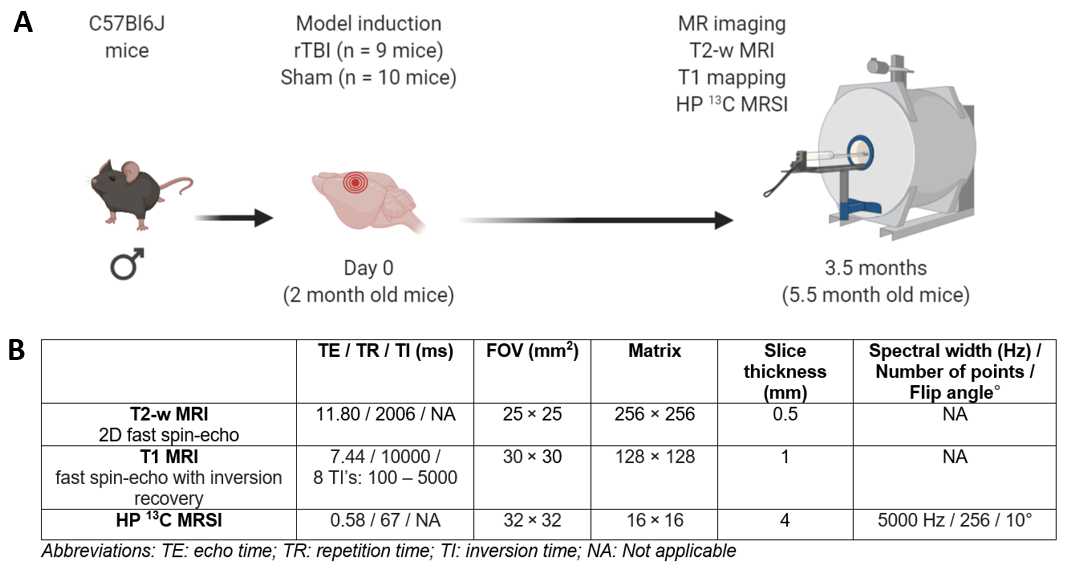
(A) Experimental timeline of the study.
Two-month old male mice received a rTBI using the CHIMERA device (n = 9 mice) or
underwent a Sham procedure (no impact, n = 10 mice). MRI was performed 3.5
months after Sham or rTBI. (B) Table summarizes parameters used for MR
acquisitions.
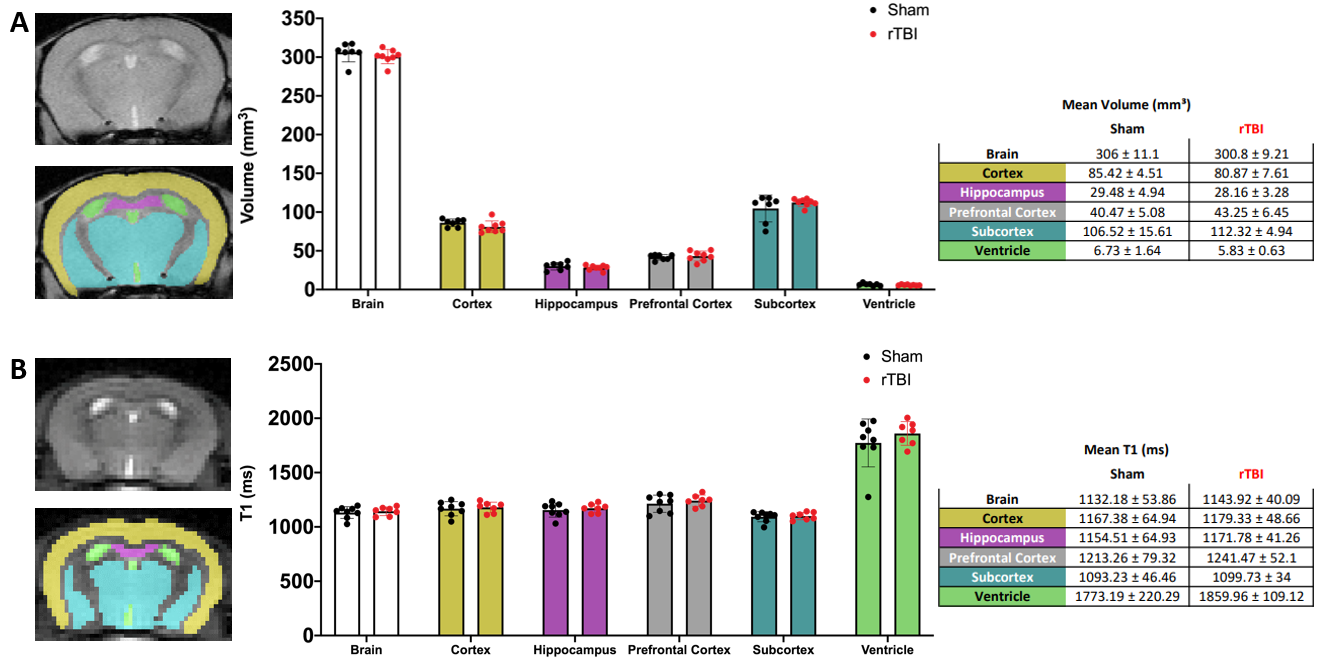
(A) Representative manual brain masking of T2-weighted MRI data and corresponding volume sizes in mm3 for Sham (n = 7) and rTBI (n = 8). Color coding for regional masks: Yellow = cortex, pink = hippocampus, gray = prefrontal cortex (not pictured), blue = subcortex, and green = ventricle. (B) Manual segmentation of regions on T1 map 128x128 data; and corresponding T1 relaxation values for Sham (n = 7) and rTBI (n = 7). All values reported as mean±std.
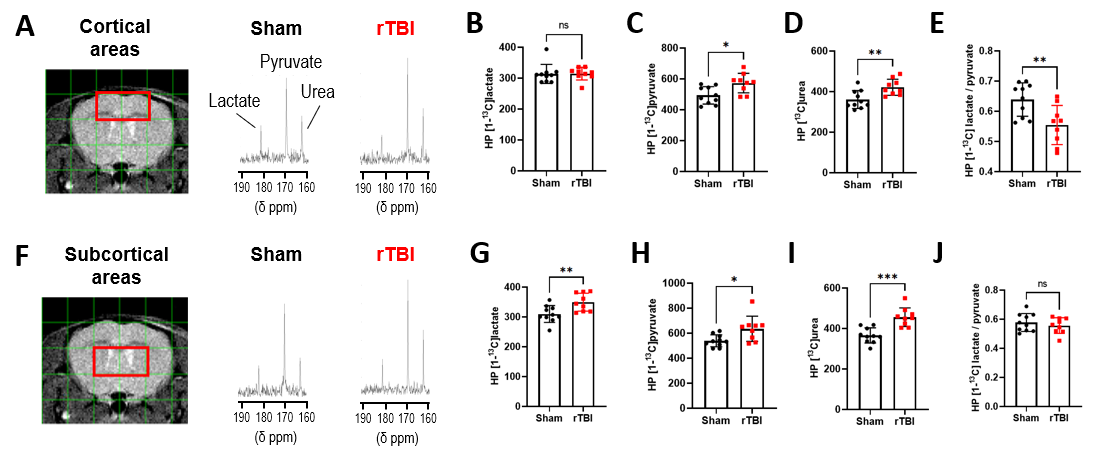
(A) T2 image and overlaid grid
used for HP 13C MRSI analyses, red voxels indicate cortical areas
and their corresponding HP 13C spectra for Sham and rTBI.
Quantitative analyses of HP 13C (B) lactate, (C) pyruvate,
(D) urea and (E) lactate/pyruvate for cortical areas. (F) T2
weighted image and HP 13C grid, red voxels indicate subcortical
areas and corresponding HP 13C spectra. Quantitative analyses of HP 13C
(G) lactate, (H) pyruvate, (I) urea and (J)
lactate/pyruvate for subcortical areas.
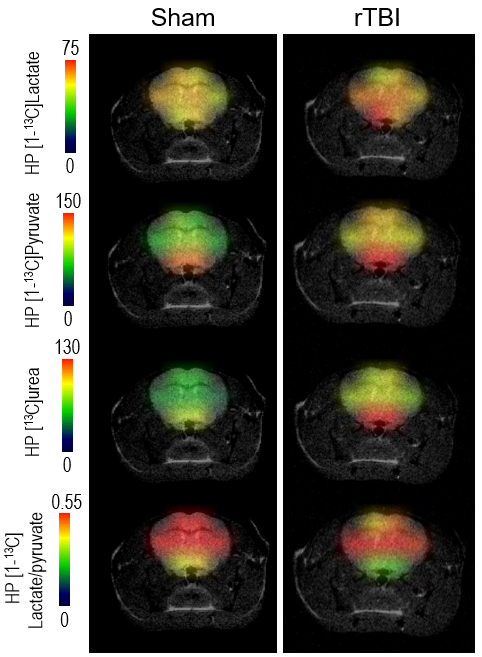
Representative
HP metabolites heatmaps from a Sham and a rTBI mouse showing higher [1-13C]lactate
levels in rTBI in subcortical areas. Higher HP [1-13C]pyruvate and
HP 13C urea levels can be observed in the entire brain of rTBI
compared to Sham. Heatmaps of HP [1-13C]lactate/pyruvate ratio show
lower values in cortical areas in rTBI.
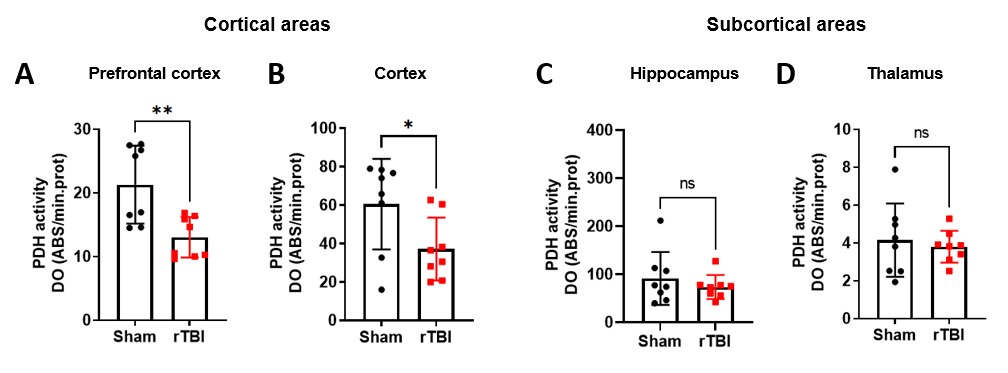
PDH
activity was decreased in cortical areas, including the (A) prefrontal
cortex and (B) cortex, of rTBI compared to Sham. PDH activity was not
significantly different in subcortical areas, including the (C)
hippocampus and (D) thalamus, between rTBI and Sham.