3978
Decreases in functional connectivity of white matter in a resting state and during a working memory task in schizophrenia1Vanderbilt University Institute of Imaging Science, Vanderbilt University Medical Center, Nashville, TN, United States, 2Biomedical Engineering, Vanderbilt University, Nashville, TN, United States, 3Radiology and Radiological Sciences, Vanderbilt University Medical Center, Nashville, TN, United States, 4Psychiatry and Behavioral Sciences, Vanderbilt University Medical Center, Nashville, TN, United States, 5Electrical Engineering and Computer Science, Vanderbilt University, Nashville, TN, United States
Synopsis
White matter (WM) pathological changes play a role in disturbing neural connectivity of schizophrenic subjects. We extended our previous analyses of WM-GM connectivity to quantify WM functional differences during a resting state and a working memory task between schizophrenic patients and healthy controls. Significant deficits of functional connectivity were found in several WM tracts in schizophrenic patients relative to healthy controls. These findings add further evidence of the presence of WM changes in schizophrenia subjects compared to controls and further illustrate the potential relevance of functional signals arising from WM in a task and at rest.
Introduction
Schizophrenia is characterized by deficits in cognitive functions including working memory1. Pathological changes in white matter (WM) have been indicated as playing a role in disturbing neural connectivity in brains of schizophrenic subjects2. MRI studies examining anatomical alterations in WM have been reported3, but possible functional deficits in WM in schizophrenia have not been evaluated.Although blood-oxygenation-level-dependent (BOLD) signals are smaller in WM than GM, BOLD fluctuations in a resting state in WM share common features with those from GM and correlate significantly with BOLD signals from specific GM areas to which they connect4-6.
In this study, we extend our previous analyses of WM-GM connectivity4,7 to quantify differences of WM fMRI metrics during a resting state and a working memory task between patients with schizophrenia and healthy controls.
Methods
Images of 84 healthy controls (CON) and 67 patients with schizophrenia (SCZ), schizoaffective disorder and schizophreniform disorder, matched for age, gender and parental education, were analyzed.Data acquisition, task design and data preprocessing
Resting state fMRI volumes (EPI, TR/TE=2s/35ms, resolution=3x3x3mm3, matrix=80x80x38, dynamics per run=300, #runs=1), task fMRI (same parameters except dynamics per run=152 and #runs=6) and T1w TFE (TR/TE=8ms/3.7ms, resolution=1x1x1mm3, matrix=256x256x170) data were acquired on a Philips 3T Achieve-DX scanner using a 32-channel head coil.
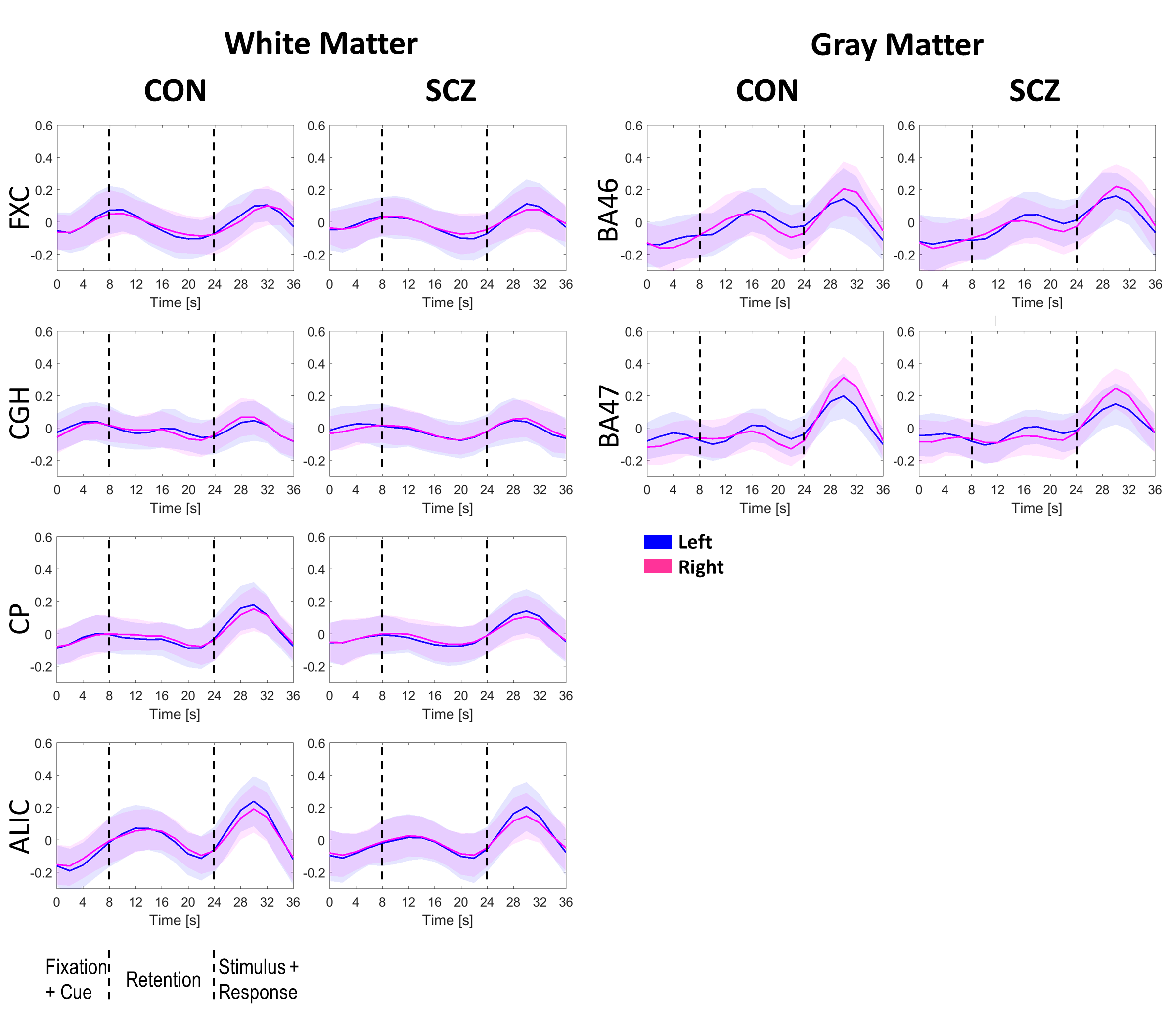
During each run of task fMRI, subjects were instructed to remember positions of objects in five memory trials and perform non-memory-related tasks in three control trials. Each memory trial consisted of a 4s fixation, a 4s encoding period, a 16s retention, a 1s stimulus and a 13s inter-trial interval that included a response.
Preprocessing of fMRI data included correcting slice timing and head motion, regressing-out 24 motion parameters and mean CSF signal, filtering (passband=0.01-0.1Hz), co-registering to the MNI space, detrending, and voxel-wise normalization of the time-courses into zero mean and unit variance. Preprocessing the T1w images included segmenting WM, GM, and cerebrospinal fluid and registering the tissue probability maps to the MNI space.
Calculation of functional correlation matrix (FCM)
The calculations of inter-regional correlations for each subject were restricted to WM and GM regions of interest (ROIs) that were defined by the Eve atlas8 (46 WM tracts) and PickAtlas9 (82 Brodmann areas) that were further constrained within masks generated by thresholding the WM and GM probability maps at 0.8. The preprocessed time-courses were averaged over the voxels within each ROI and for each pair of WM and GM ROIs they were then cross correlated, excluding any time points with large motions (frame-wise displacement10 >0.5). The resulting 46x82 correlation coefficients formed an FCM of WM-GM pairs (FCMWG). The possible influences of gender, race, age, maternal and paternal years of education were regressed out from FCMWG using a generalized linear model.
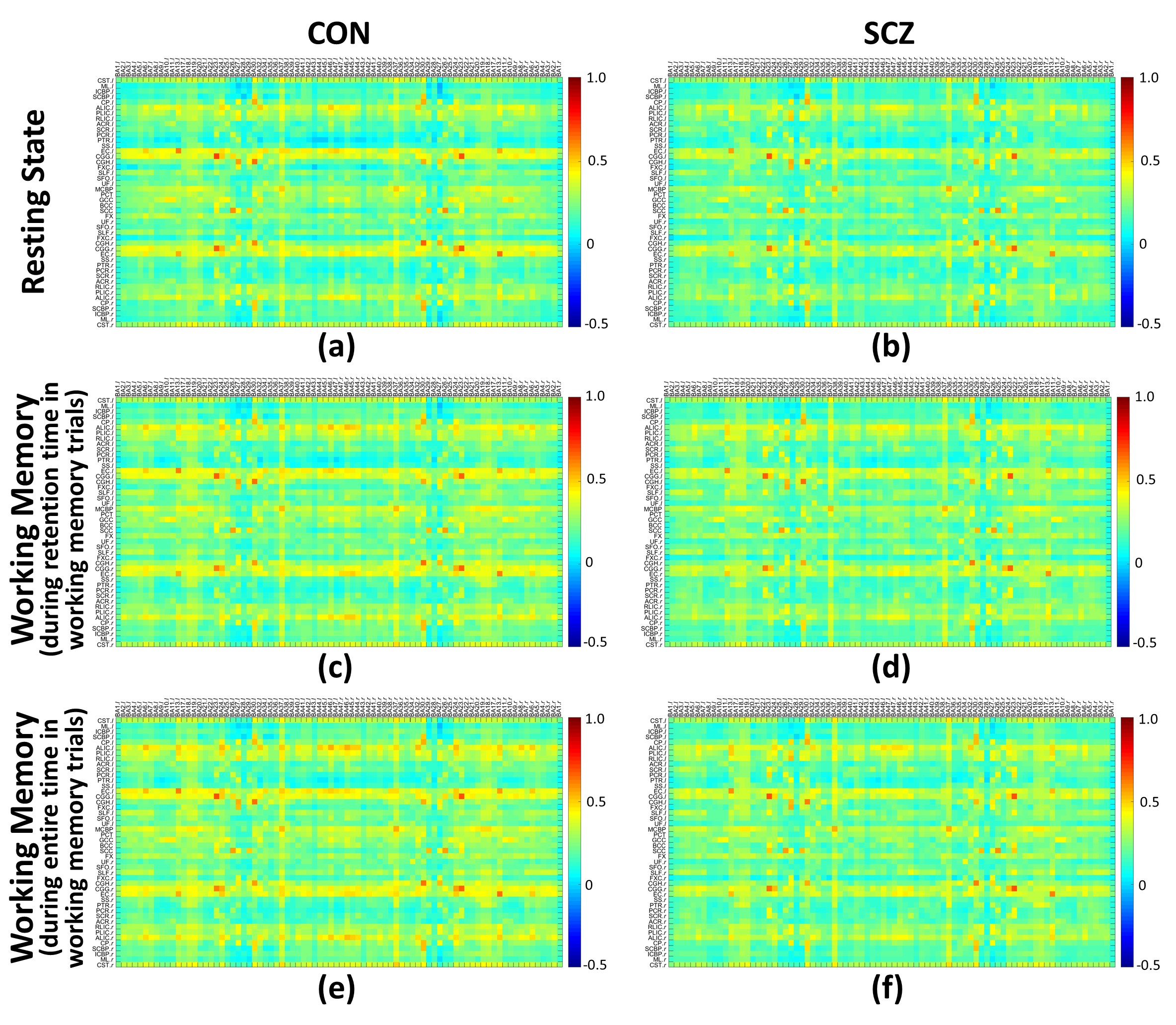
Each subject had three FCMWG calculated using the time-courses for three specific conditions. In the resting state scenario, each time-course included all 300 dynamics. In the first working memory scenario, each time-course included these dynamics during the retention time of working memory trials. In the second working memory scenario, each time-course included the dynamics during entire memory trials.
Statistical analysis
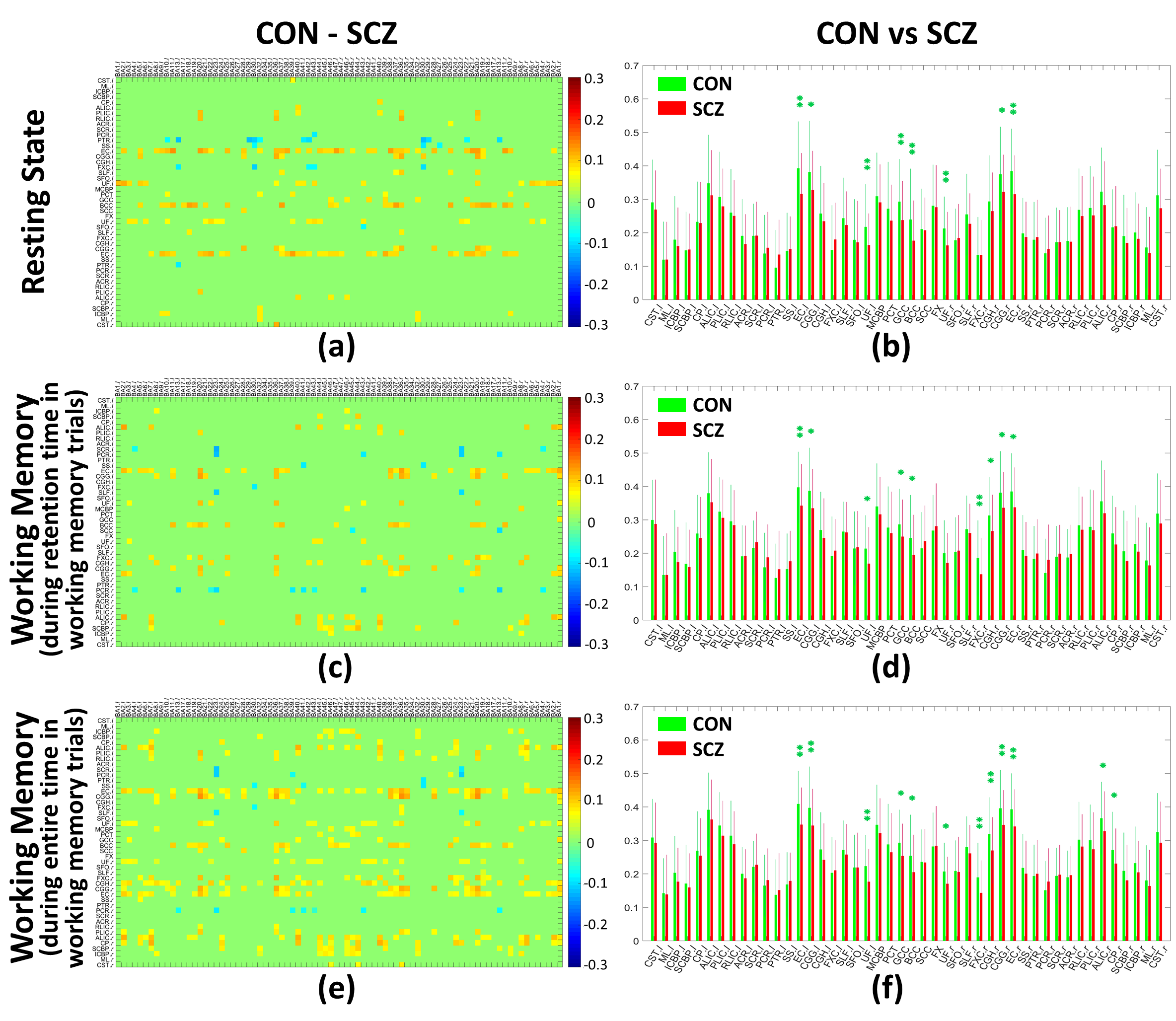
For each scenario, the FCMWG across subjects within each group were averaged and the group differences in the mean FCMWG values were calculated. Unpaired t-tests were conducted for each FCMWG element across subjects within the two groups. The resulting P-values were corrected for multiple comparisons using a false discovery rate11, PFDR. To estimate the overall connectivity of each WM tract, the FCMWG elements corresponding to each WM ROI were averaged. The mean and standard deviation of each WM-tract-wise connectivity across subjects within each group were then calculated. Values of the WM-tract-wise connectivity in the SCZ group were compared with the CON group using unpaired-sample t-tests.
Results and Discussions
The group means of FCMWG for the CON and SCZ groups in the three scenarios are shown in Figure 1 (Table 1 shows the names of all WM and GM ROIs in FCMWG).Figure 2(a,c,e) shows the differences in the mean FCMWG values between groups under three scenarios. In a resting state (Figure 2(b)), significant decreases were found in external capsule (ECl,r), cingulum (cingulate gyrus) (CGGl,r), uncinate fasciculus (UFl,r), genu and body of corpus callosum (GCC and BCC) in the SCZ group relative to the CON group. These WM tracts were previously reported to be different in schizophrenia12-15 using structural MRI. In the first memory scenario (Figure 2(d)), significant decreases were measured in ECl,r, CGGl,r, UFl, GCC, BCC, fornix (FXCr) and cingulum (hippocampus) (CGHr). In the second memory scenario (Figure 2(f)), significant decreases were also found in UFr, anterior limb of internal capsule (ALICr) and cerebral peduncle (CPr). Compared to resting state, the memory scenarios identified functional differences in FXCr and CGHr, tracts which are believed to be engaged in spatial memory tasks16,17. The decreases in ALICr and CPr, found in only the second memory scenario, may be related to the onset of the stimulus response (Figure 3). These findings add further evidence of the presence of WM changes in SCZ subjects compared to CON subjects and also further illustrate the potential relevance of functional signals arising from WM in a task and at rest.
* Gore and Woodward contributed equally.
Acknowledgements
This work was supported by NIH grant NS093669 (Gore), MH102266 (Woodward) and Vanderbilt Discovery Grant FF600670 (Gao). We also thank Advanced Computing Center for Research and Education (ACCRE) at Vanderbilt University for distributed computation.References
1. Park S, Holzman PS. Schizophrenics Show Spatial Working Memory Deficits. JAMA Psychiatry. 1992;49(12):975-982.
2. Davis KL, Stewart DG, Friedman JI, et al. White Matter Changes in Schizophrenia: Evidence for Myelin-Related Dysfunction. JAMA Psychiatry. 2003;60(5):443-456.
3. Nestor PG, Kubicki M, Gurrera RJ, et al. Neuropsychological correlates of diffusion tensor imaging in schizophrenia. Neuropsychology. 2004;18(4):629-637.
4. Ding Z, Huang Y, Bailey SK, et al. Detection of synchronous brain activity in white matter tracts at rest and under functional loading. Proceedings of the National Academy of Sciences. 2018;115(3):595-600.
5. Gore JC, Li M, Gao Y, et al. Functional MRI and resting state connectivity in white matter - a mini-review. Magnetic Resonance Imaging. 2019;63:1-11.
6. Li M, Newton AT, Anderson AW, Ding Z, Gore JC. Characterization of the hemodynamic response function in white matter tracts for event-related fMRI. Nature Communications. 2019;10(1):1140. 7. Gao Y, Li M, Zu Z, et al. Progressive degeneration of white matter functional connectivity in Alzheimer’s Disease. SPIE Medical Imaging; 2019; San Diego, California, United States.
8. Oishi K, Faria A, Jiang H, et al. Atlas-based whole brain white matter analysis using large deformation diffeomorphic metric mapping: Application to normal elderly and Alzheimer's disease participants. NeuroImage. 2009;46(2):486-499.
9. Lancaster JL, Woldorff MG, Parsons LM, et al. Automated Talairach Atlas labels for functional brain mapping. Human Brain Mapping. 2000;10(3):120-131.
10. Power JD, Barnes KA, Snyder AZ, Schlaggar BL, Petersen SE. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage. 2012;59(3):2142-2154.
11. Benjamini Y, Hochberg Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. Journal of the Royal Statistical Society Series B (Methodological). 1995;57(1):289-300.
12. Lee S-H, Kubicki M, Asami T, et al. Extensive white matter abnormalities in patients with first-episode schizophrenia: A diffusion tensor imaging (DTI) study. Schizophrenia Research. 2013;143(2):231-238.
13. Kubicki M, Westin C-F, Nestor PG, et al. Cingulate fasciculus integrity disruption in schizophrenia: a magnetic resonance diffusion tensor imaging study. Biological Psychiatry. 2003;54(11):1171-1180.
14. McIntosh AM, Maniega SM, Lymer GKS, et al. White Matter Tractography in Bipolar Disorder and Schizophrenia. Biological Psychiatry. 2008;64(12):1088-1092.
15. Skudlarski P, Schretlen DJ, Thaker GK, et al. Diffusion Tensor Imaging White Matter Endophenotypes in Patients With Schizophrenia or Psychotic Bipolar Disorder and Their Relatives. American Journal of Psychiatry. 2013;170(8):886-898.
16. Gaffan D. Dissociated effects of perirhinal cortex ablation, fornix transection and amygdalectomy: evidence for multiple memory systems in the primate temporal lobe. Experimental Brain Research. 1994;99(3):411-422.
17. Parker A, Gaffan D. The effect of anterior thalamic and cingulate cortex lesions on object-in-place memory in monkeys. Neuropsychologia. 1997;35(8):1093-1102.
Figures