2948
Downfield Magnetic Resonance Spectroscopy in a mouse model of Brain Glioma
Sónia Isabel Gonçalves1, Rui V. Simões1, and Noam Shemesh1
1Champalimaud Research, Champalimaud Centre for the Unknown, Lisbon, Portugal
1Champalimaud Research, Champalimaud Centre for the Unknown, Lisbon, Portugal
Synopsis
MRS allows non-invasive in-vivo exploration of tissue metabolism. However, about half of the proton spectrum (downfield of water) has been nearly ignored over the decades of MRS application due to water suppression. We show that ISIS-based Relaxation Enhanced MRS (iRE-MRS) which uses frequency selective excitation and ISIS localization offers short echo times and enhances exchange-broadened resonances. For the first time, we measure in-vivo downfield spectra in mouse glioma tumors (and controls) and show remarkable spectral signatures for the tumor downfield.
Introduction
Downfield magnetic resonance spectroscopy (MRS) has received little attention insofar due to the difficulty in detecting resonances mostly originating from labile protons. Only a few studies with high dynamic range MRS [1-5] or Relaxation-Enhanced MRS [6] (RE-MRS), focused on downfield spectra [7, 8], and revealed (i) interesting resonances undetectable in the upfield counterpart (NAD NADH) [9] and (ii) exchange-related phenomena [5]. A recently introduced method, termed iRE-MRS [1], combines RE-MRS [6] with Image-Selected-In-Vivo-Spectroscopy (ISIS) localization [10] in the aim of obtaining downfield spectra at TEs as short as 5 ms. Given the rapidly exchanging signals downfield, we hypothesized that short TE downfield spectra may provide insight in disease in general and in tumors in particular, where changes in environment may lead to differences in metabolic concentrations but also perhaps exchange properties. Here, we use iRE-MRS to investigate, for the first time, in-vivo downfield MRS spectra in glioma tumors in-vivo.Methods
Animal experiments were preapproved by the institutional and national authorities and carried out according to European Directive 2010/63. Experiments were carried out on wild type mice, N=4 control (4 males, mean weight = 21.2±1.3 g) and N=5 tumors (2 males, mean weight = 20.9±1.3 g), aged ~5-6 months. Animal preparation. Tumors were induced in all N=5 animals by intracranial stereotactic injection of 10e5 GL261 cells in the caudate nucleus as reported previously in [11]. Pulse sequence design. Figure 1 illustrates the iRE-MRS pulse sequence. It is based on spectrally-selective excitation and refocusing, avoiding water suppression [5]. However, unlike the previous RE-MRS studies, localization is achieved by an ISIS block, consisting of three spatially-selective adiabatic inversions. OVS modules were implemented to improve localization quality. Spectrally-selective RF pulses. The spectrally-selective RF pulses were generated by the shape algorithm implemented in Paravision 6.0.1 (Bruker Biospin, Ettlingen, Germany), which employs the Shinar LeRoux algorithm [12]. Excitation and refocusing pulses were centered at 9.5 ppm, having a bandwidth of 6.5 ppm. MRS experiments. All experiments took place approximately 2.5 weeks after injection and were performed using a 9.4 T horizontal bore scanner (Bruker-Biospin, Karlsruhe, Germany), equipped with an 86 mm coil for transmission and a 4-element array cryocoil (Bruker BioSpin, Fallanden, Switzerland) for signal reception. iRE-MRS spectra were obtained in a 2.2×2.2×2.2 mm3 voxel, located in the striatum. Experiments were executed using the following common acquisition parameters: TR=15000 ms (>>5T1 of metabolites), 8 steps per ISIS cycle, 15 repetitions (cycles) per experiment, spectral width 5597.01 Hz, spectral resolution 1.94Hz/point. Post-processing. Each FID (ISIS cycle) was Fourier transformed, the resulting spectra were individually rephased and then averaged over the total number of repetitions for each scan. Apodization was carried-out with an exponential factor of 20 Hz-width. All spectra were scaled to the amplitude of the unsuppressed water peak and aligned with respect to the ‘NAA’ amide peak falling at approximately 7.8 ppm [13].Results
Figure 2 shows the voxel positioning and downfield spectrum in one illustrative animal. Figure 3 shows individual overlaid raw iRE-MRS spectra for controls (Figure 3a) and tumors (Figure 3b). Visible differences between controls and tumors can be observed in all spectral peaks, especially with respect to peak amplitude and to some extent, also alignment. Figure 4 shows the average spectra spectrum over animals for both controls (Figure 4a) and tumors (Figure 4b). The average spectrum for controls closely reproduces previous results in the rat [14], with peaks labeled as “NAA”, a, b, c, d and e easily distinguishable. Other smaller peaks, located at ~8.5 (f), 8.25 (g), and 9.0 (j), 9.3 (i), and 9.5 (h) ppm are also apparent. For tumors, the average spectrum shows evident differences: strong attenuation of peaks “NAA”, b, c, and e, while peaks j, i, and h completely disappear.Discussion
Our results indicate that in-vivo downfield spectra of glioma tumors are sensitive to the disease through marked spectral changes. Tumor growth involves a complex interplay of factors [15-17] that comprise, amongst others, disturbed metabolism, and acidification of the cellular environment which can influence the exchange rates and peak locations of exchanging protons [8]. Our findings already suggest quite a few interesting targets for future research: (1) Peaks j, i and h, which probably correspond to the NAD+ proton resonances [9], completely disappear in tumor, suggesting redox potential changes in the tumor area [18]; (2) We observed peak location shifts which may represent pH effects [8]. Such hypothesis can be tested in the future by employing water-suppression and observing how exchange impacts these signals directly [6]. Finally, though more research is required in terms of spectral assignment, the changes observed here may differ in different types of tumors. All these vistas augur well for downfield MRS as a sensitive marker for cancer.Conclusion
In-vivo downfield iRE-MRS of glioma tumors revealed marked spectral differences indicating features other than those typically associated with upfield counterparts, such as changes in redox potential and pH. These findings strongly motivate future studies exploring downfield MRS signatures.Acknowledgements
The work of Rui V. Simões was funded by grant H2020-MSCA-IF-2018, ref 844776.References
[1] Gonçalves et al., ISMRM 2018; [2] Piotto et al., J. Biomol. NMR, 2, 661-665, 1992; [3] Dreher and Leibfritz, Magn. Res. Med., 54, 190-195, 2005. [4] MacMillan et al., 65, 1239-1246, 2011; [5] MacMillan et al., 70, 916-924, 2013; [6] Shemesh et al., Nat Commun, doi: 10.1038/ncomms5958, 2014; [7] van Zijl and Moonen, Magn. Res. Med., 29: 381-385, 1993; [8] Mori et al., Magn. Res. Med., 40, 36-42, 1998; [9] Graaf and Behar Magn. Res. Med., 27, 802-809, 2014; [10] Ordidge et al., J. Magn. Res., 66, 283, 1986; [11] Simões et al., NMR Biomed, 21, 251-264, 2008; [12] Pauly et al., IEEE Trans. Med. Imaging, 10, 53-65, 1991; [13] Fichtner at al., Magn. Res. Med., 78, 11-19, 2017 [14] Gonçalves et al., Magn. Res. Med., 82(4), 1266-77, 2019; [15] Korenjak and Zavadil, Cancer Sci., Oct 8, 2019; [16] Hsieh and Tsai, World J Gastrointest Oncol, 11(9): 686-704, 2019; [17] Markopoulos et al., Cells, 8(19), 2019; [18] Acharya et al., Oxid Med Cell Longev, 3(1), 23-34, 2010.Figures
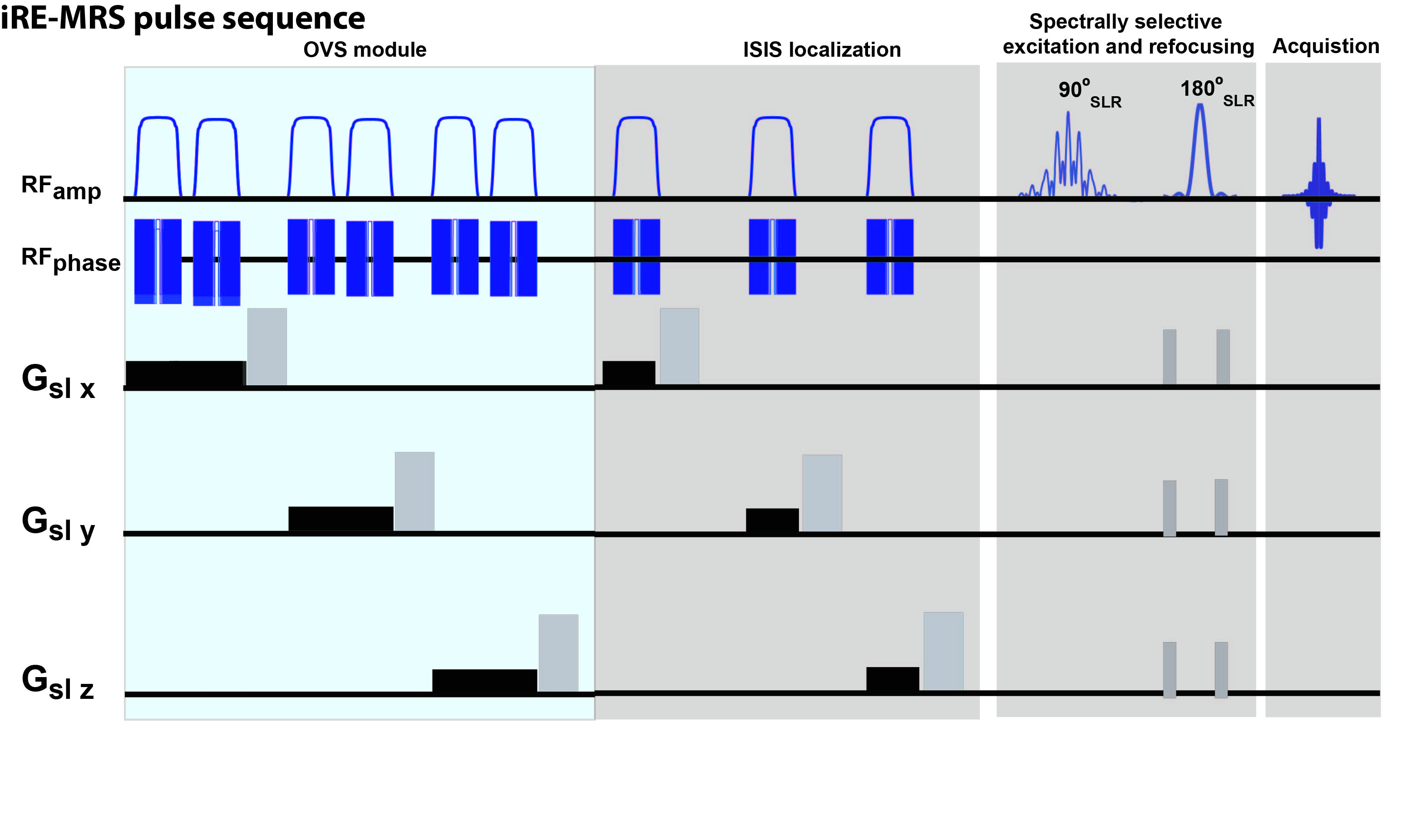
Figure 1. Pulse sequence
diagram.
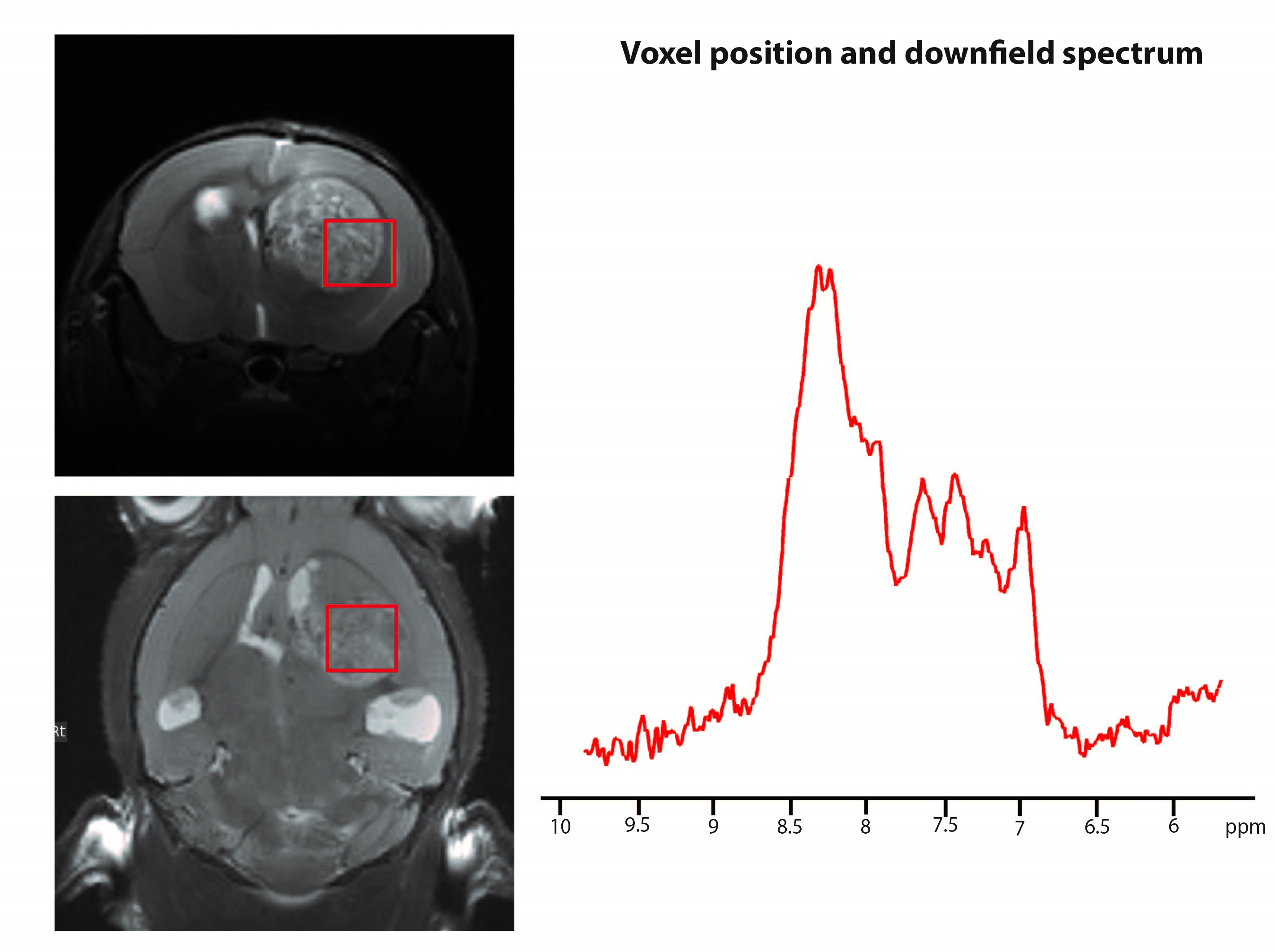
Figure 2. Voxel position and
corresponding downfield spectrum in an illustrative animal.
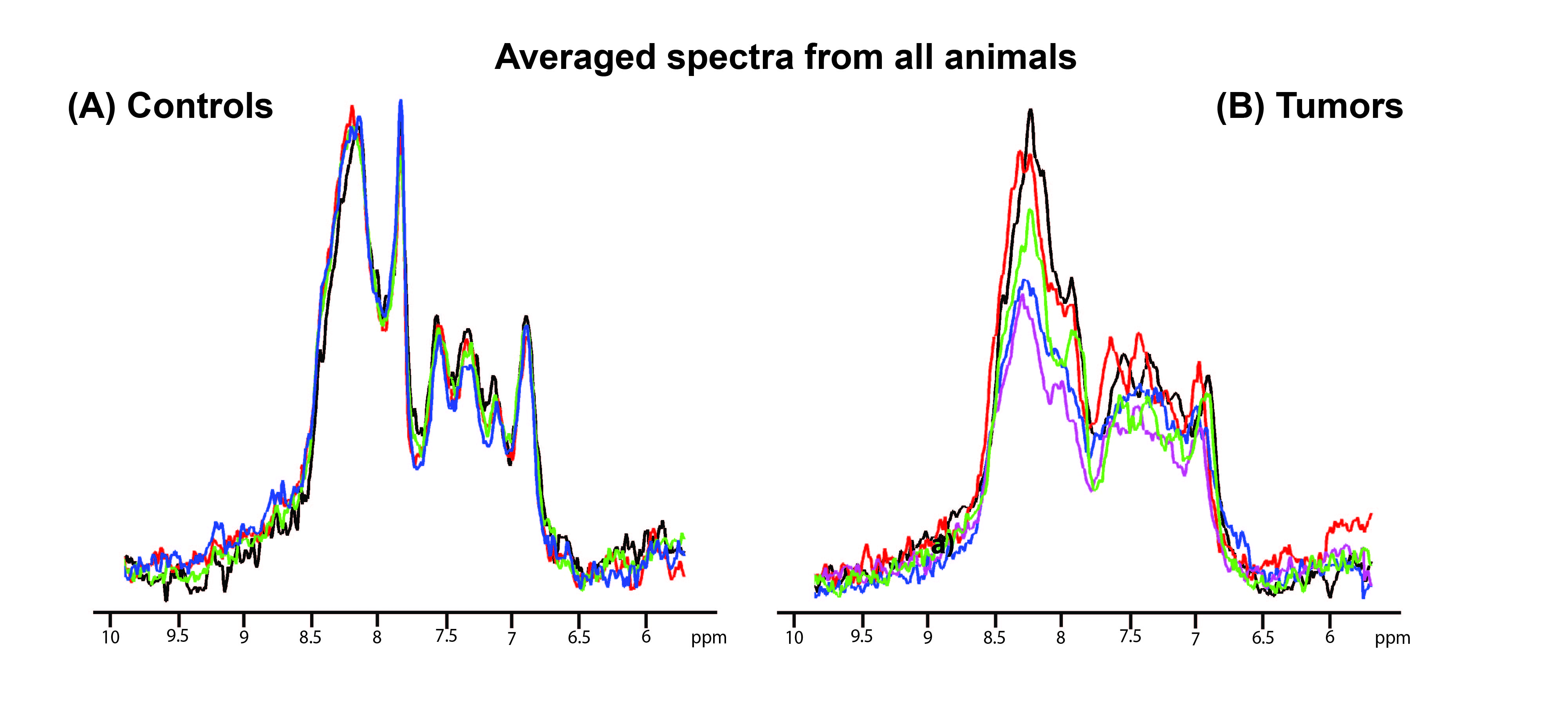
Figure 3. Overlaid spectra of
all animals. a) Controls b) Tumors.
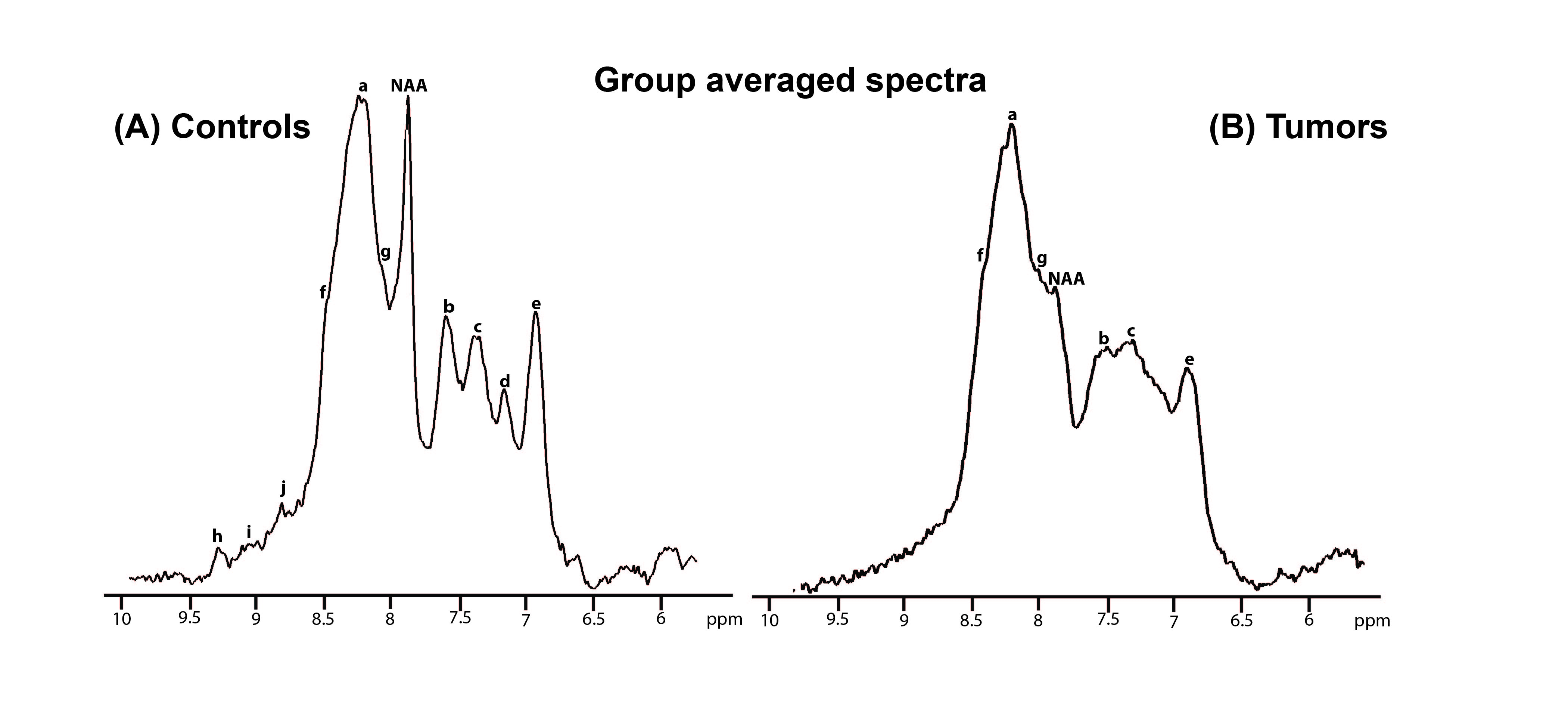
Figure 4. Group averaged spectra.
a) Controls b) Tumors.