2858
J-refocused semi-LASER spectroscopy at 7T: proof-of-concept and application in the human brain at 7T
Chloé Najac1, Vincent Boer2, Hermien E. Kan1, Andrew G. Webb1, and Itamar Ronen1
1C.J. Gorter Center for High Field MRI, Department of Radiology, Leiden University Medical Center, Leiden, Netherlands, 2Danish Research Centre for Magnetic Resonance, Copenhagen University Hospital Hvidovre, Copenhagen, Denmark
1C.J. Gorter Center for High Field MRI, Department of Radiology, Leiden University Medical Center, Leiden, Netherlands, 2Danish Research Centre for Magnetic Resonance, Copenhagen University Hospital Hvidovre, Copenhagen, Denmark
Synopsis
Spin-echo sequences suffer from loss of signal due to J-modulation for coupled spins. Previously, it was shown that adding a π/2 pulse between two π pulses in a double spin-echo sequence partially refocuses some of the J-evolution. However, the potential of such sequence at ultrahigh field is limited due to the large chemical shift displacement error. Here, we propose a J-refocused variant of the sLASER sequence (J-sLASER) to improve quantification of J-coupled metabolites at ultrahigh field. Significant improvement in quantitation of J-coupled metabolites is illustrated using simulation, phantom and in vivo measurements in the human brain.
Introduction
Spin-echo sequences suffer from loss of signal due to J-modulation for coupled spins in metabolites of interest such as glutamate, lactate, glutamine and others. Although shorter TE values improve signal-to-noise ratios (SNR) and limits loss due to J-modulation, it also results in significant confounding contributions from macromolecules (MM). It was shown that an additional π/2 RF pulse between two π pulses in a double spin-echo sequence refocuses some of the dephasing caused by J-evolution1,2. This has been successfully implemented in the PRESS sequence, reducing much of the signal loss due to homonuclear J-coupling3-6. At ultrahigh field, poor localization accuracy due to large chemical shift displacement error (CSDE) in the PRESS sequence remains problematic7. Additionally, the increase in linewidth, typically about 10Hz and above, without concomitant increase in J-splitting results in a greater degree of signal cancellation due to anti-phase peaks cancellation. Here, we propose a J-refocused variant of the semi-LASER (sLASER) sequence, which provides both accurate localization and significant recovery of signal from J-coupled systems. The sequence (J-sLASER) was first validated using simulation followed by phantom acquisition at 7T. Finally, the sequence was compared to sLASER sequence in the human brain, illustrating significant improvement in J-coupled metabolites quantification using J-sLASER.Materials and methods
J-sLASER sequence: The classical sLASER sequence was modified to incorporate a slice-selective π/2 RF pulse and concomitant slice selection and refocusing gradients at time of formation of the first echo (Figure 1). The phase of the added π/2 pulse was set to be orthogonal to the first π/2 pulse. Phase cycling was used to suppress unwanted signal due to potential out-of-volume excitation, pulse angle deviations or T1 relaxation in the first half of the sequence. All experiments were conducted on a Philips 7T whole body MRI scanner (Philips Healthcare, The Netherlands) equipped with a volume transmit/32-channel receive head coil (Nova Medical, USA).Simulation: Resonances of protons belonging to metabolites detectable in the human brain were simulated using Matlab programs based on the density matrix formalism and published chemical shifts and J-coupling values8. The simulation did not include T2 relaxation. Signal was integrated using Matlab and signal J-modulations as a function of TE (up to 500ms) for both J-sLASER and sLASER sequences were investigated.
Phantom validation: In vitro sequence validation was subsequently performed on a GE-MR spectroscopy “Braino” phantom using both sequences and a range of TE values from 44 to 164ms and with TR=4000ms, number of signal averaged (NSA)=8, bandwidth=3kHz, 2048 data points. Spectra were corrected for eddy currents using unsuppressed water spectra. Metabolites signal was quantified using Matlab and LCModel9. A basis-set for each TE value was created for both sequences.
In vivo measurement: In vivo experiments were performed on 3 healthy volunteers. A 3D-T1W gradient-echo acquisition (TR/TE=5/2ms, resolution 1x1x1mm) was used for planning of the experiments. A 8mL volume-of-interest was positioned in the posterior cingulate cortex (PCC, Figure 4). Metabolites and water spectra were acquired using both sLASER and J-sLASER sequence at TE=45 and 65ms (TR=5600ms/NSA=80/BW=3kH/np=1024). Spectra were corrected for eddy currents using an in-house Matlab routine and quantified using LCModel9. A basis-set including MM spectrum was created for both sequences and TE values.
Results and discussion
Figure 2 shows results of spectral simulations for some of the J-coupled proton groups in brain metabolites for classic sLASER and J-sLASER sequences over a range of TE values. This illustrates the potential to improve detection of metabolites such as lactate, glutamate and glutamine at intermediate TE values.Figure 3 shows phantom results obtained with both sequences. It can be seen that the evolution of the J-modulation for glutamate and lactate follows the theoretical prediction in the simulation (Figure 2B). A slight decrease in the NAA singlet signal with J-sLASER is due the generation of residual stimulated echoes caused by the additional π/2 pulse and the slice selection gradient.
In vivo measurements were performed in the PCC of the human brain (n=3). Figure 4 shows representative spectra acquired with J-sLASER and sLASER at TE=45ms. The dramatic effect of the J-refocusing is seen on all glutamate peaks, as well as on the lactate and glutamine. Finally, the CRLB values for three metabolites obtained with LCModel from data acquired with both sequences at two different TE values are reported in Table 1. While the overall SNR was slightly lower due to loss of signal by the stimulated echo generated by the added π/2 pulse, the CRLB decreased for glutamate and glutamine at both TE values and for lactate at the shorter TE with the J-sLASER sequence.
Conclusion
We illustrated the potential of the J-sLASER sequence to improve detection of J-coupled resonances in the human brain at ultrahigh field. We found in vivo a strong improvement in glutamate detection. Further investigations are needed to improve SNR, for example by using a non-selective π/2 pulse and thus minimizing the stimulated echo generated by it, and aim to decrease TE below 45ms. The J-sLASER sequence offers benefits that may be used to improve the measurement of small changes in metabolites in functional MRS experiments10,11, as well as new perspective on spectral editing, by combining data from sLASER and J-sLASER in conditions that optimally maximize the differences for desired J-coupling systems.Acknowledgements
The authors would like to thank Drs. D. Deelchand and P.G. Henry from Center of Magnetic Resonance Research at University of Minnesota (USA) and Dr. Julien Valette at Atomic Energy and Alternative Energies Commission in Paris (France) for sharing their Matlab programs to create LCModel basis-sets.References
[1] P. van Zijl et al. J. Magn. Res. 1990; [2] K. Takegoshi et al. J. Magn. Res. 1989; [3] Pan et al., MRM 1996; [4] Pan et al., MRM 2010; [5] Mulkern et al., MRM 1996; [6] Chen et al., NM in Biomed. 2013; [7] D.K. Deelchand et al., MRM 2017; [8] V. Govindaraju et al., NMR in Biomed. 2000; [9] Provencher et al., MRM 1993; [10] Schaller et al., NeuroImage 2014; [11] Bednařík et al., J. Cereb. Blood Flow Metab. (2015)Figures
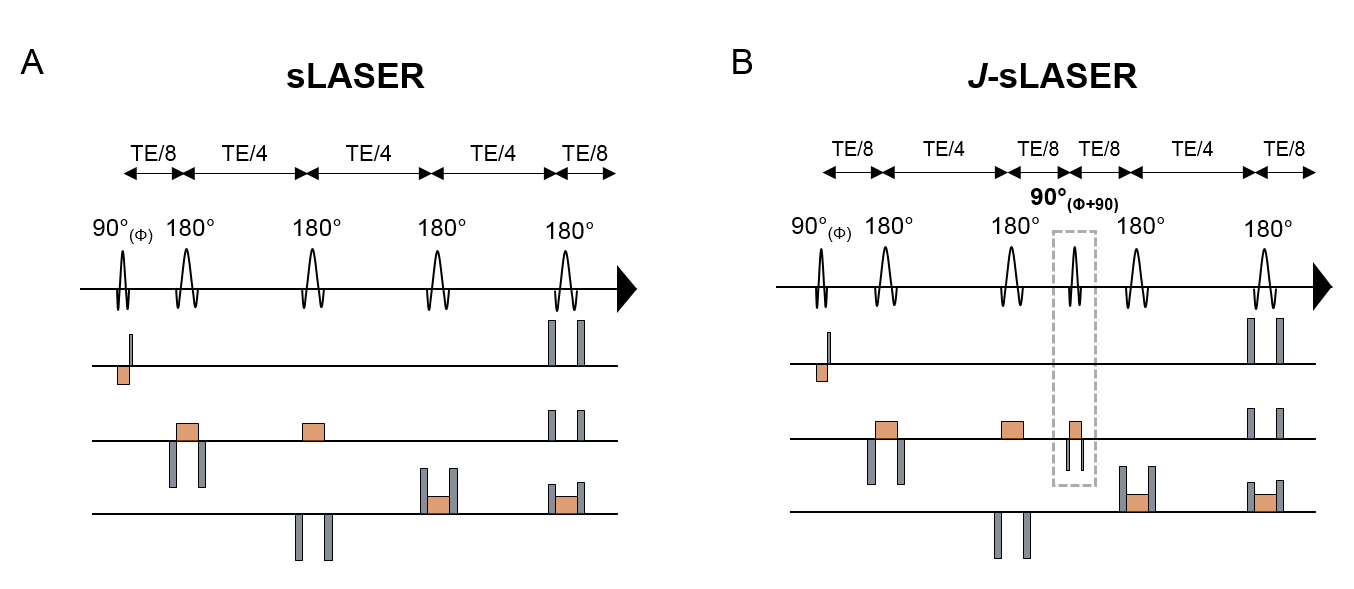
Figure 1: The
(A) sLASER and (B) J-sLASER sequences implemented
and used in this study. Slice selection gradients are represented in orange and
refocusing/spoiler gradients are shown in grey. The additional π/2 RF pulse and concomitant slice selection and refocusing
gradients in J-sLASER sequence are highlighted using the
dotted rectangle.
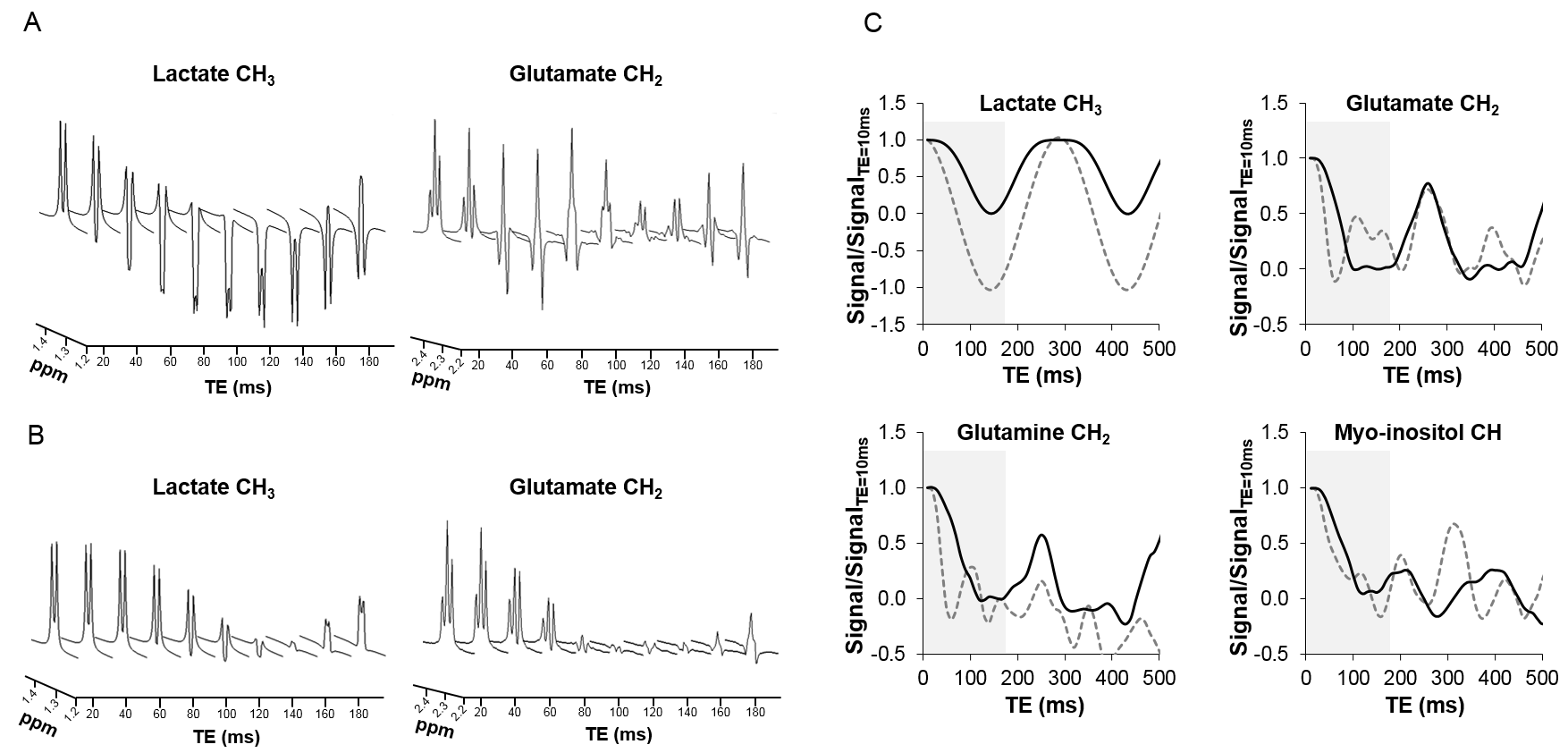
Figure 2: (A) Simulated spectra of lactate and
glutamate resonances as a function of TE for sLASER and (B) J-sLASER sequences; (C) glutamate, lactate, glutamine and myo-inositol signals over a
range of TE values as measured with sLASER (grey dotted line) and J-sLASER (black solid line). This
illustrates the strong potential of J-sLASER
to improve detection of J-coupled protons.
The grey area is used to highlight the range of TE values investigated in
phantom (Figure 3).
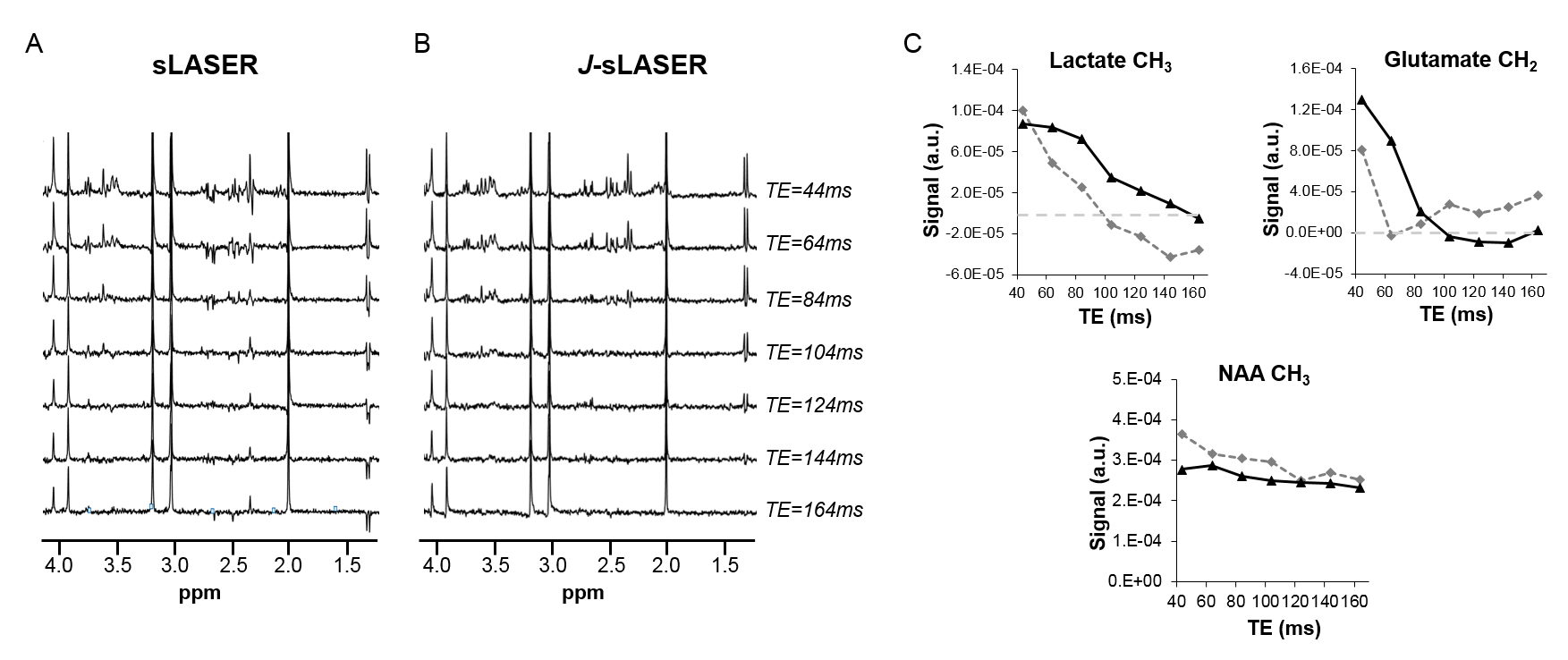
Figure 3: (A) Representative in vitro spectra acquired with sLASER and (B) J-sLASER sequences and
for different TEs are shown (44, 64, 84, 104, 124, 144, 164ms). Linewidth in
phantom was below 2Hz, resulting in nice lactate detection at all TE
using sLASER. In vivo due to broader
linewidth such contribution would not be detectable due to cancellation of anti-phase peaks. (C) Quantification of lactate, glutamate and NAA signals are
reported for both sLASER (grey dotted line, diamond) and J-sLASER (black solid line, triangle) sequences.
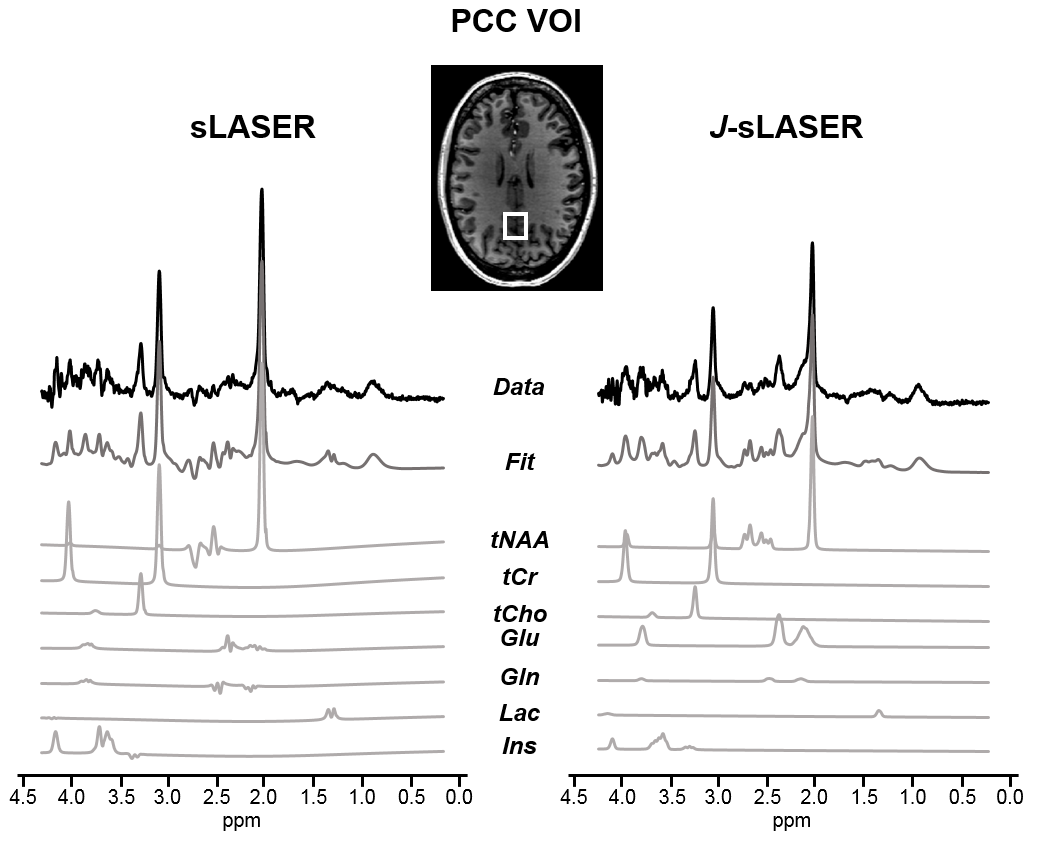
Figure 4: In vivo spectra acquired in PCC region in one healthy volunteer with both sequences:
sLASER (left) and J-sLASER (right).
The LCModel fits are shown below. Note the dramatic increase in glutamate
peaks, also in 2.1ppm and 3.7ppm. LCModel decomposition is shown for few
metabolites included in the basis-set: tNAA (total N-acetyl-aspartate), tCr
(total creatine), tCho (total choline), glutamate (Glu), glutamine (Gln),
lactate (Lac) and myo-inositol (Ins).
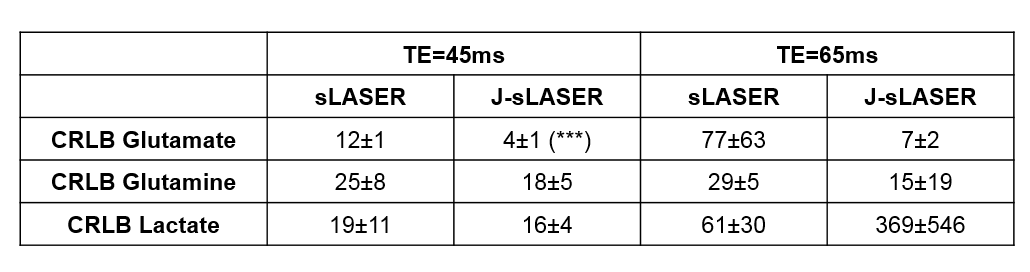
Table 1: CRLB
values (mean±s.d.) for three metabolites and for both sequences and both TE
values based on the in vivo data. A
Student's t-test was used to compare
sLASER and J-sLASER showing
significant decrease in CRLB for glutamate at TE=45ms.