0631
Cerebral metabolism of hyperpolarized [2H7, U-13C6]D-glucose in the healthy mouse under different anesthetic conditions1LIFMET, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland, 2Danmarks Tekniske Universitet, Lyngby, Denmark, 3CIBM, Ecole Polytechnique Fédérale de Lausanne, Lausanne, Switzerland
Synopsis
Glucose is the primary fuel for the brain and its metabolism is linked with cerebral function. Isoflurane anesthesia is commonly employed in preclinical MRS but influences functional connectivity. The combination of isoflurane and medetomidine is regularly used in rodent fMRI and show similar functional connectivity as in awake animals. Here we compared the cerebral metabolism of hyperpolarized [2H7,U-13C6]-D-glucose under these two anesthetic conditions. When using the combination, the [1-13C]lactate signal and lactate-to-glucose ratio were more than doubled compared to isoflurane solely, showing that the change of anesthesia had a high impact on cerebral glucose uptake and glycolytic flux.
Introduction
Cerebral function is associated with high metabolic activity and requires a continuous supply of oxygen and nutrients from the bloodstream. The mammalian brain is mainly supported by glucose as fuel to meet its high metabolic demand1. Under physiological conditions, glucose metabolism is tightly linked with cerebral function2,3.Hyperpolarized (HP) MR of endogenous molecules by dissolution dynamic nuclear polarization (dDNP)4 is a powerful method for studying intermediary metabolism in real-time5. It was demonstrated that HP [2H7, U-13C6]-D-glucose can be measured in vivo6 and employed for the real time detection of glycolytic reactions in a lymphoma mouse model7 and in the healthy mouse brain8.
Isoflurane is one of the most common anesthetic drug used in preclinical MRS metabolic studies, but is known to affect cerebral hemodynamics9,10 and to influence functional connectivity11. In the context of HP 13C MRS, isoflurane was shown to alter the cerebral metabolism of HP pyruvate12-14. The combination of isoflurane and medetomidine is routinely employed in fMRI in rodents, and it enables to maintain similar functional connectivity as in the awake animal11,15. Given that glucose metabolism is linked to neuronal activity2, the aim of this study was to compare the cerebral metabolism of HP [2H7, U-13C6]-D-glucose under these two anesthetic conditions.
Methods
MR measurements were performed in a 9.4T MRI system(Varian/Magnex) using a home-built quadrature 1H-coil/13C surface coil that was placed on top of the mouse head. Brain metabolism of HP glucose was monitored in two groups of 12h-fasted male C57BL6/J mice (N=14). In the first group (ISO), mice were kept under isoflurane for the entire duration of the experiment (1.3-1.6% isoflurane; N=8). In the second group (MED+ISO), anesthesia was switched to a combination of medetomidine and isoflurane one hour before injection of HP [2H7, U-13C6]D-glucose (medetomidine 0.3 mg/kg bolus followed by 0.03 mg/kg/h i.v. combined with 0.25-0.5% isoflurane; N=6). Single voxel 1H MRS measurements were carried out in the hippocampus (2x2.8x2mm3) of each mouse 10 minutes before the HP [2H7, U-13C6]D-glucose bolus. Animal physiology was constantly monitored and complete blood analysis were performed before the mouse entered the scanner (1.25h before HP bolus) and after the end of the 13C MRS acquisition (˜2min post bolus).HP glucose was prepared as previously described16, and polarized in a 7T/1K DNP polarizer17, yielding liquid-state polarization of 29±4%. To monitor real-time de novo synthesis of [1-13C]lactate, a 540μL of 44±10mM HP [2H7,13C6]D-glucose was injected through the femoral vein. A series of pulse-acquire sequence was triggered 5.5s post injection with 25° frequency selective Gaussian pulse (250μs) centered at 182 ppm every 1s for 70s. Lactate-to-glucose ratio (LGR) was calculated from the summed spectra.
Results
Typical dynamic spectra shows that in both anesthetic conditions, the injection of HP [2H7, U-13C6]D-glucose results in de novo synthesis of [1-13C]lactate. The timecourses of all animals in both groups were well reproducible (Fig.1).For the two groups, the initial blood glycemia, the post bolus blood glycemia, and 13C glucose fractional enrichment were kept similar as those may influence the metabolism of HP [2H7, U-13C6]D-glucose (Fig.2).
The total LGR was found significantly higher in the MED+ISO group compared to ISO group (+128%, Fig.3A) and the [1-13C]lactate signal was detected for a longer period of time in the MED+ISO group (Fig.1).
Steady-state metabolites concentrations deduced from 1H MRS spectra showed significantly higher lactate pool-size and creatine-to-phosphocreatine ratio (Cr/PCr) in animals anesthetized by isoflurane (Fig.4). All other metabolites in the neurochemical profile did not show any significant difference.
Discussion
Here we compared the cerebral metabolism of HP glucose between animals anesthetized with isoflurane and a combination of isoflurane and medetomidine. The production of [1-13C]lactate from [2H7,13C6]D-glucose is a consequence of 12 enzymatic steps including glucose transport, glycolysis and pyruvate conversion to lactate. We found that the amount of [1-13C]lactate and LGRs were more than doubled in animals anesthetized with the combination than with isoflurane solely. The Cr/PCr ratio that is related to ATP and ADP balance was found significantly different between the two groups, indicating a difference in the energetic state. Additionally, endogenous lactate concentration was higher in the ISO group, implying that [1-13C]lactate production from HP glucose is not a reflection of the steady-state pool-size but rather related to the dynamic of the glycolytic flux.Isoflurane at clinical concentration (1.4-2.5%) significantly reduces glucose transport18 and local cerebral glucose utilization in all cortical areas19. HP [1-13C]lactate production depends on HP glucose uptake and on the metabolic demands. As the mouse cortex was well positioned under the sensitive area of the coil, the cortical areas may bring the main contribution the observed differences. The correlation between isoflurane concentration and LGR supports this hypothesis, implying that little variations in isoflurane concentration directly influence the dynamic of lactate synthesis (Fig.3B). Interestingly, a larger amount of lactate was produced in the case when the functional connectivity was similar to the awake animals (MED+ISO)11. This is in line with several studies suggesting that astrocytic lactate production can be stimulated during neuronal activation20-23.
Conclusion
We report that change of anesthesia had high impact on cerebral glucose uptake and glycolytic flux. The higher SNR reported in this work is an important step toward real-time imaging of cerebral glycolysis.Acknowledgements
Supported by CIBM of the UNIL, UNIGE, HUG, CHUV, EPFL, the Leenaards and Jeantet Foundations.References
1. Mergenthaler, P. et al., Trends Neurosci, 2013
2. Sonnay, S. et al., Front Neurosci., 2017
3. Sokoloff L., Neurochem Res., 1999
4. Ardenkjaer-Larsen, J. H. et al.,Proc Natl Acad Sci USA, 2003
5. Comment, A. & Merritt, M. E., Biochemistry, 2014
6. Allouche-Arnon et al., Contrast Media & Molecular Imaging, 2012
7. Rodrigues, T. B. et al. Nat. Med., 2014
8. Mishkovsky, M. et al., Scientific Reports, 2017
9. Todd, M. et al., J Neurosurg Anesthesiol.,1996
10. Conzen, P. et al., Anesth Analg.,1992
11. Paasonen, J. et al., NeuroImage., 2018
12. Josan, S. et al., MRM, 2012
13. Hypponen V. et al., ISMRM 2019
14. Marjanska et al., NMR in biomedecine, 2018
15. Grandjean, J. et al., NeuroImage, 2014
16. Capozzi, A. et al., J. Phys. Chem. Lett, 2019
17. Cheng, T. et al, NMR in biomedicine, 2013
18. Andersen et al. Anesthesiology, 1973
19. Ori, C. et al., Anesthesiology, 1986
20. Hertz, L. et al., Neurochemical Research, 1988
21. Magistretti, P.J. et al., Dev. Neurosci., 1993
22. Tsacopoulos, M., Magistretti P.J. et al., The journal of Neurosci., 1996
23. Prichard, J. et al, Proc. Natl. Acad. Sci, 1991
24. Provencher, S. W, Magnetic resonance in medicine, 1993.
Figures
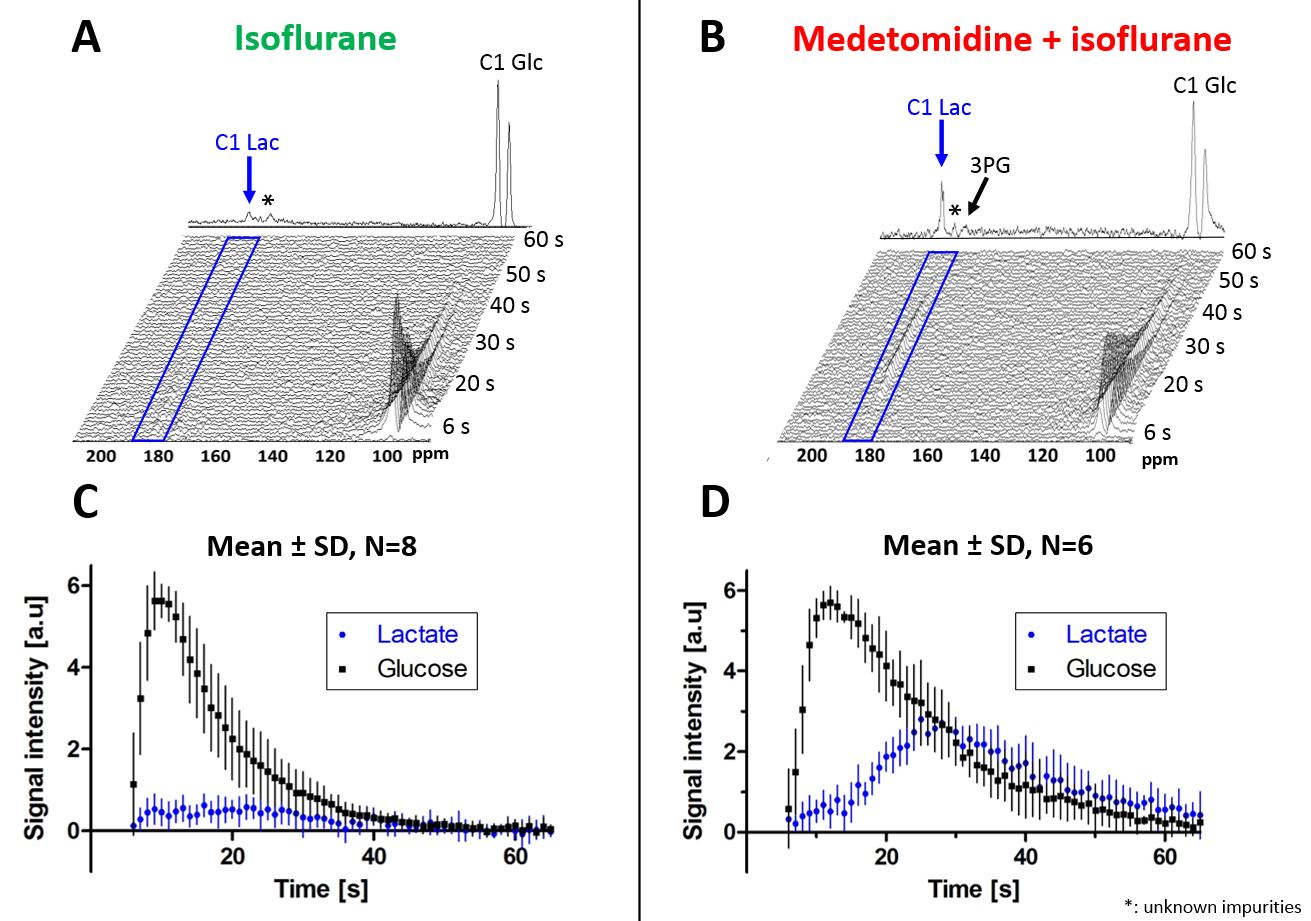
Fig.1
A,B - Characteristic spectra measured in a mouse brain under isoflurane (ISO) and under medetomidine + isoflurane (MED+ISO) anesthesia following infusion of HP [2H7,13C6]D-glucose. The glycolytic intermediate 3PG (179.8ppm) can be identified in the summed spectra in addition to the lactate at 183.5ppm. The broad peak at 175 ppm designated by (*) is an impurity in the [2H7,13C6]D-glucose powder.
C,D - The corresponding timecourses (mean ± SD) of [1-13C]lactate and [2H7,13C6]D-glucose are shown for each group
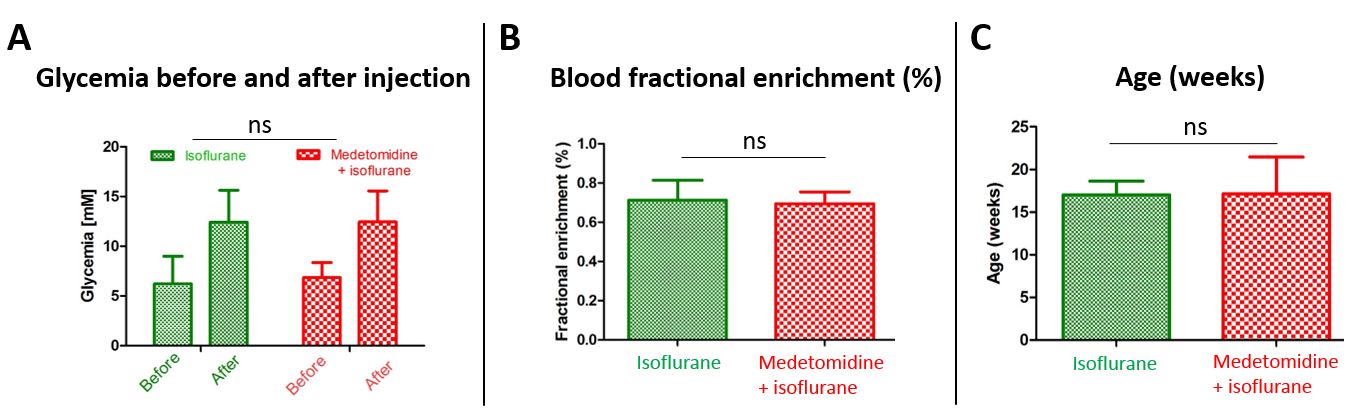
Fig.2
A – Glycemia measured when the mice enter the scanner (‘before’) and 2 minutes after injection of HP [2H7,13C6]D-glucose (‘after’) in the two anesthesia groups.
B, C - No significant differences in 13C blood fractional enrichment (B) or age (C) between the two anesthesia groups.
Statistical analysis was performed using two-way or one-way ANOVA in GraphPad Prism software.
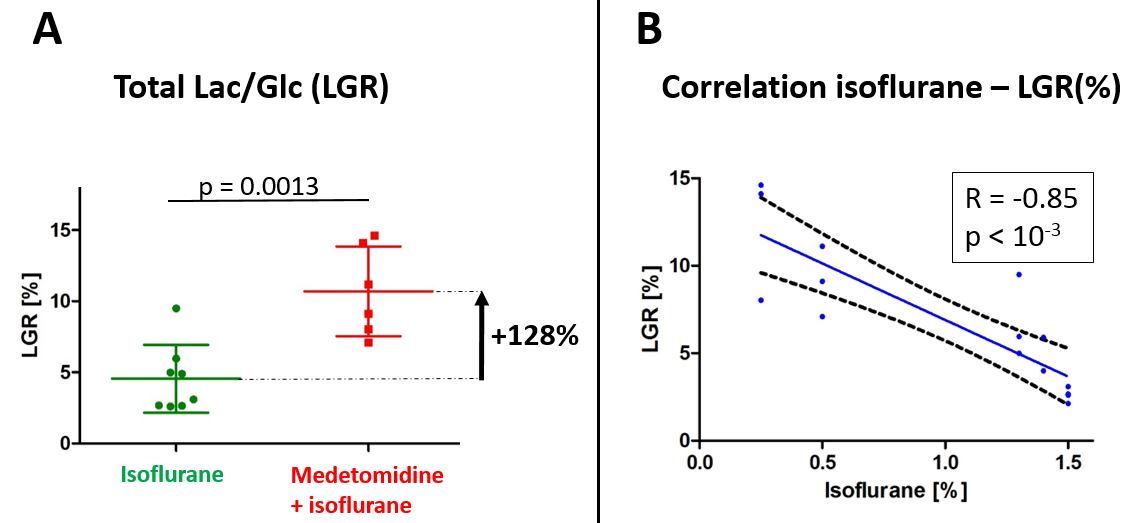
Fig.3
A - Comparison of LGR between isoflurane (ISO) and medetomidine + isoflurane (MED+ISO) groups.
B – Correlation between LGR and isoflurane concentration(%).
Statistical analysis was performed using one-way ANOVA in GraphPad Prism software.
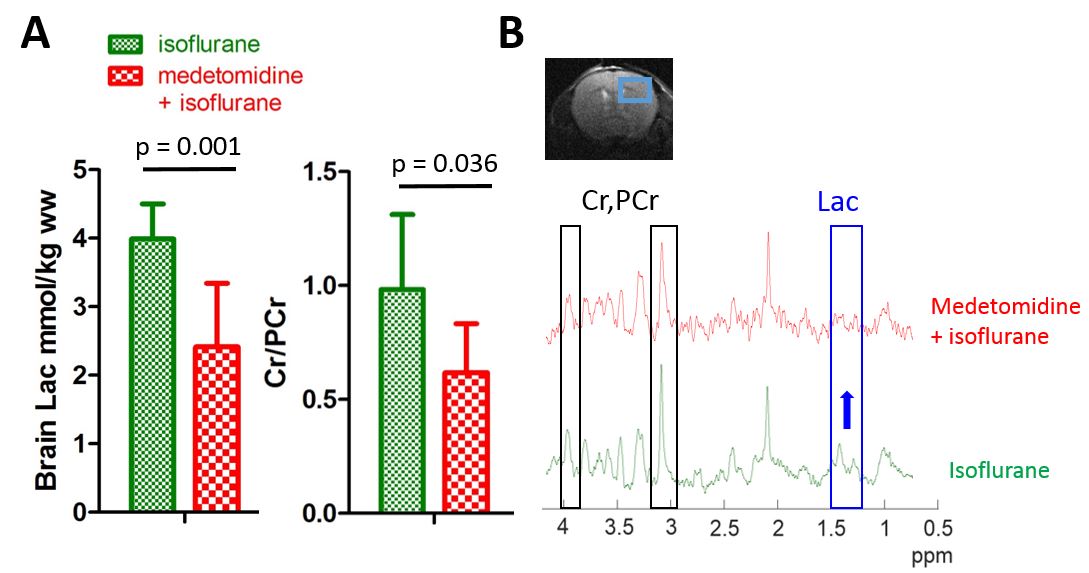
Fig.4
A - Brain Lac and Cr/PCr comparison (mean±SD) between the ISO group and the MED+ISO group.
B - Representative 1H MRS spectra in the hippocampus of an ISO mouse (green) and a MED+ISO mouse (red). Part of the spectra corresponding to Lac and Cr&PCr are shown in blue and dark, respectively.
Metabolite concentrations were estimated by LCModel using water as internal reference24 and one-way ANOVA statistical tests were performed using GraphPad Prism software.