0375
Myocardial Pi/PCr and pH during stress at 7T with STEAM 31P MRS in dilated cardiomyopathy; heart failure beyond the ejection fraction.1Cardiovascular Medicine, University of Oxford, Oxford, United Kingdom, 2Physiology, Anatomy and Genetics, University of Oxford, Oxford, United Kingdom, 3Physiology Anatomy and Genetics, University of Oxford, Oxford, United Kingdom, 4Oxford Centre for Magentic Resonance, University of Oxford, Oxford, United Kingdom, 5Medical University of Vienna, Vienna, Austria, 6Oxford Centre for Magnetic Resonance, University of Oxford, Oxford, United Kingdom, 7Imaging Methods, Slovak Academy of Sciences, Bratislava, Slovakia, 8Clinical Neurosciences, University of Cambridge, Cambridge, United Kingdom
Synopsis
The addition of Pi/PCr quantification adds value over PCr/ATP for the characterisation of myocardial energetics. In defining the chemical shift of the Pi resonance, pH can also be computed. Such measurements however are hampered in 31P MRS due to the overlapping 2,3-DPG resonance. In harnessing the black blood contrast offered by STEAM, we successfully characterise Pi (and hence myocardial pH) in a cohort of patients with dilated cardiomyopathy. We go on to shown that in these patients (but not controls) Pi/PCr rises significantly during dobutamine stress, a finding that would significantly impair the free energy of ATP hydrolysis during exertion.
Background:
ATP hydrolysis yields free energy (ΔGATP), required for active mechanisms to proceed, whose magnitude is determined by the phosphorylation potential ([ATP]/[ADP][Pi]). Cardiac 31P MRS is currently limited to determination of PCr/ATP due to the Pi resonance being obscured by 2,3 DPG from blood. Harnessing the properties of STEAM [1], we recently demonstrated robust Pi:PCr measurement at rest and, and no change during stress in healthy volunteers [2]. ATP homeostasis in early heart failure, alongside its falls in the later stages of disease [3] may make PCr/ATP, although shown to be depressed in NICM [4], a less sensitive marker of impaired energy metabolism. Pi however is shown to increase in compensated hypertrophy despite normal [ATP] [5], is detrimental to ΔGATP, and is a direct consequence of impaired CK flux [6]. Therefore, we aimed to measure Pi/PCr, PCr/ATP and pH (computed via the chemical shift of Pi relative to PRs) using STEAM 31P-MRS in a NICM group and compare these with a control group at rest. We then examine the response of Pi/PCr and pH to catecholamine stress in the NICM group.Method:
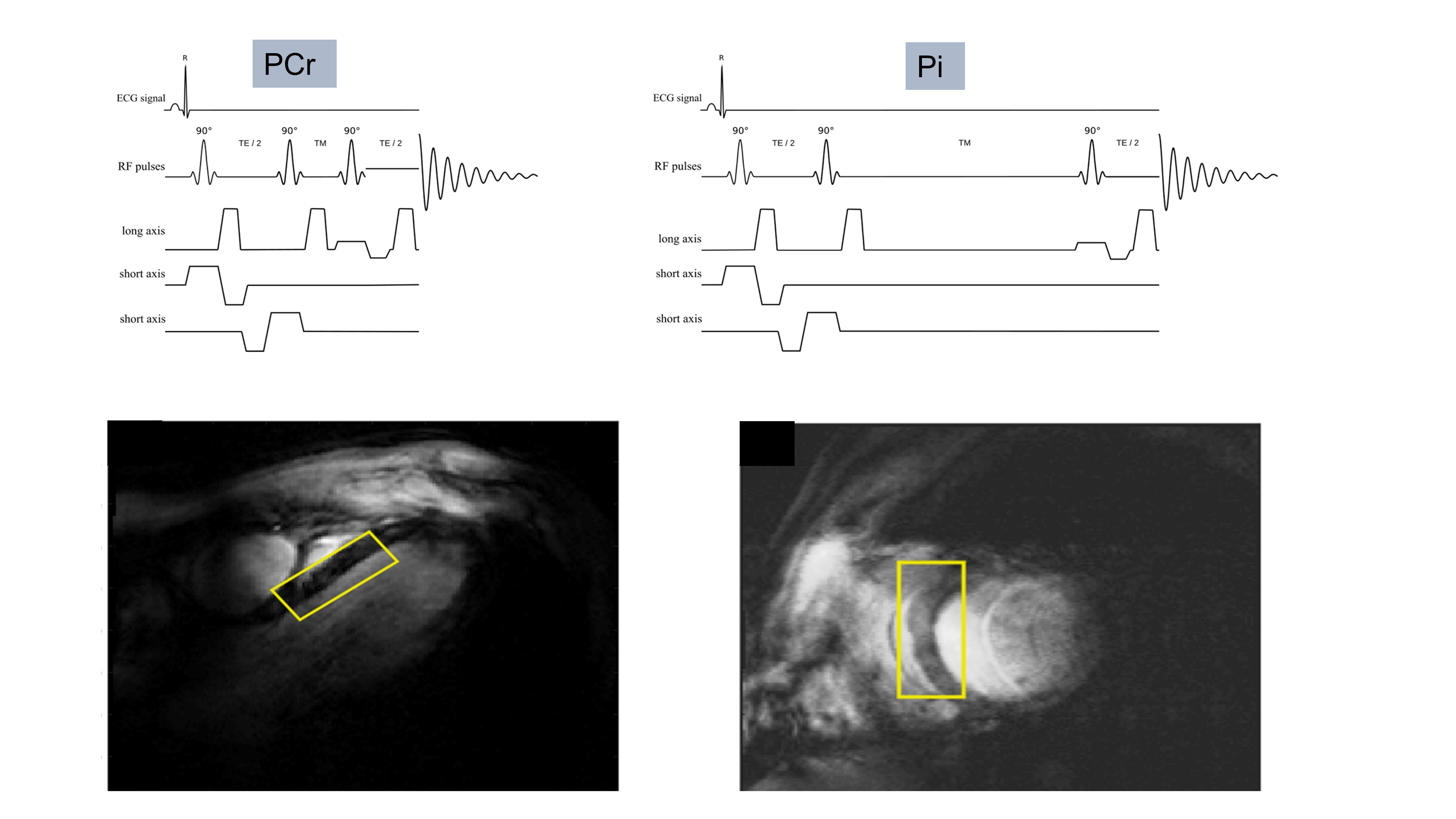
21 patients with NICM (age 58±16, BMI 25±4 kg/m2 LVEF 40±6%, LVEDVi 125±40mls/m2) all of whom had typical CMR characteristics of idiopathic DCM, and 9 age matched controls (age 49±10, BMI 24±2) were recruited. MRS was performed using a 1H loop for localisers and a 16-channel array (RAPID biomedical) for 31P MRS at a 7T MR scanner (Siemens). PCr/γ-ATP was acquired at rest with an ultra-short echo time CSI sequence (TR 2.2s, matrix size 8x16x6). Pi/PCr was acquired from the septum using interleaved STEAM acquisitions [2]. (fig 1).Results:
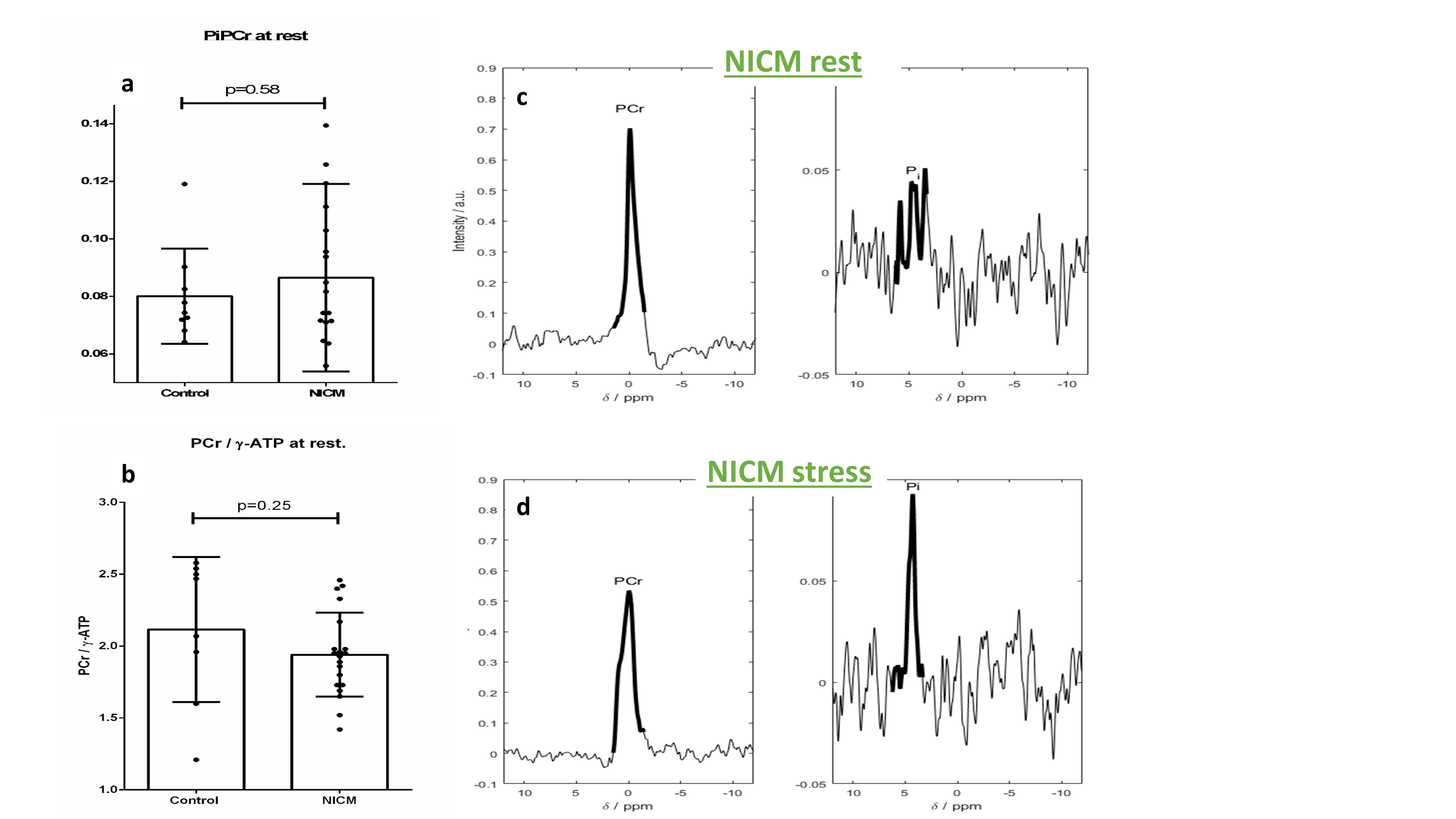
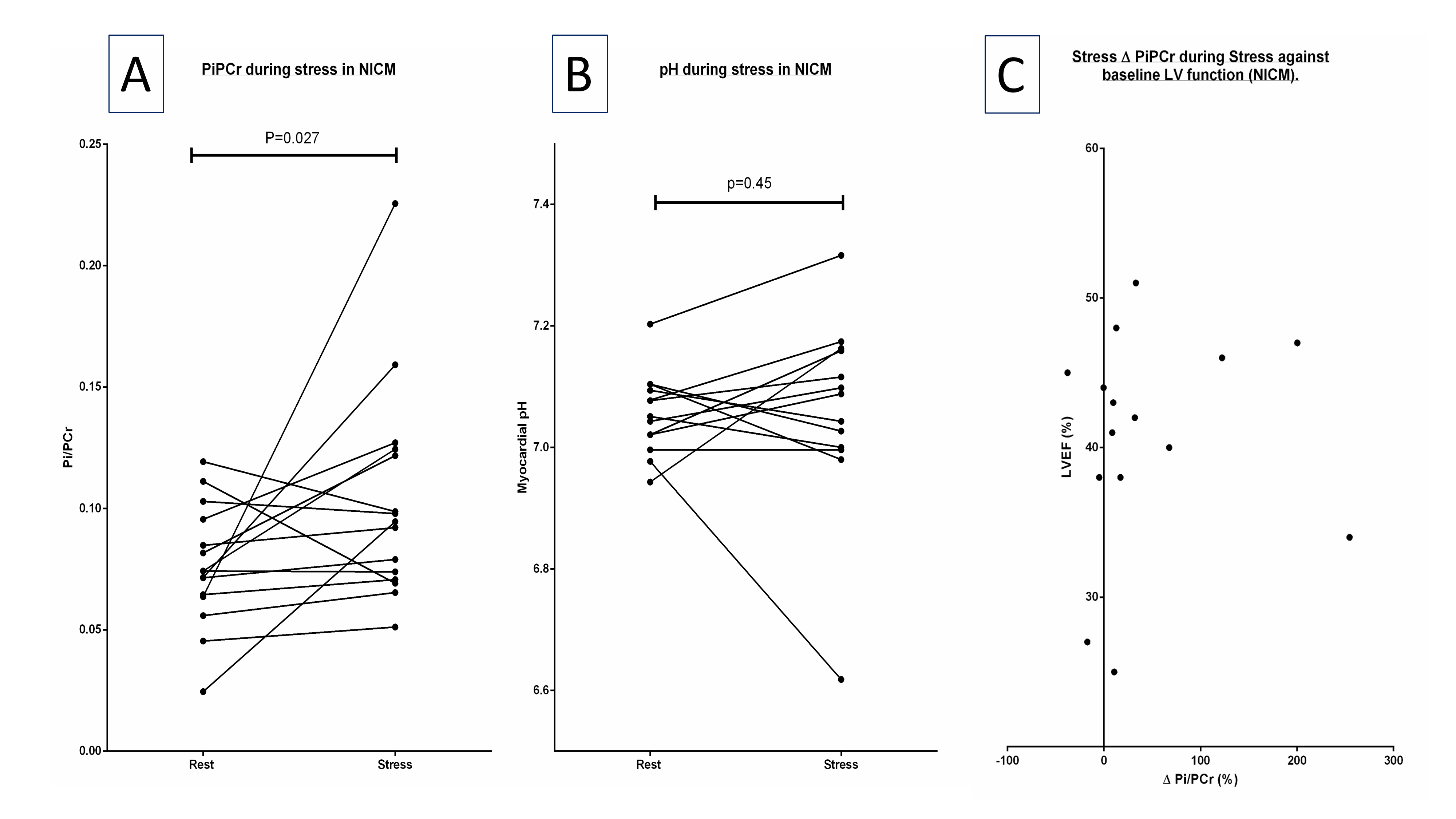
Rest Pi:PCr trended higher in the NICM group (0.087±0.033 vs. 0.080±0.017 p=0.58) with a corresponding trend lower in PCr/γ-ATP (1.94±0.29 vs. 2.12±0.50 p=0.25, fig 2a,b). Rest myocardial pH trended lower in NICM (7.02±0.11 vs. 7.08±0.11 p=0.21). 18/21 NICM patients underwent dobutamine stress (rate pressure product [mmHg*bpm] rest vs stress 7972 vs 15357 [p<0.001]) a typical pair of spectra are seen in figure 2. Pi:PCr rose significantly during stress by 39% (0.076±0.025 vs 0.103±0.044, t(14) = 2.11, p=0.03 (figure 3A), Although myocardial pH was unchanged (7.06±0.07 vs. 7.06±0.16, t(12) = 0.12, p=0.45, figure 3B). ΔPi:PCr bore no correlation to underlying LVEF (R2 = 0.005, p=0.8, figure 3C).Discussion:
In contrast to previous reports demonstrating stability during stress in controls [2], we demonstrate a significant increase in Pi during stress in NICM which bore no correlation to underlying ventricular function. This rise far exceeding that considered physiological to match ATP production (oxidative phosphorylation) with rates of hydrolysis [7], likely reflects impaired CK flux [8] with inadequate ATP regeneration exposed during times of increased demand. This rise has a deleterious impact on exertional ΔGATP and supports the notion that energetic impairment, independent of systolic function, may account for the heterogenous symptomatology experienced by this group during exertion.Acknowledgements
CR receives funding from the Sir Henry Dale Fellowship (Wellcome trust and the royal society) [098436/Z/12/B]
AS receives funding from the FWF Schrodinger Fellowship [J3032]
LV receives funding from the Slovak Grant Agencies VEGA [2/0001/17] and APVV [15-0029]
AA and MP receive funding from the British heart foundations from Clinical Research training fellowships.
References
1 Karaus A, Merboldt KD, Graessner J, et al. Black‐blood imaging of the human heart using rapid stimulated echo acquisition mode (STEAM) MRI. Journal of Magnetic Resonance Imaging: An Official Journal of the International Society for Magnetic Resonance in Medicine 2007;26:1666-71.
2 Apps A VL, Peterzan M, Neubauer S, Rogers C, Rider O, Schmid AI. Myocardial pH and Pi in healthy volunteers remain stable during moderate catecholamine stress; a 7T 31P spectroscopy study. In proceedings of the 22nd Annual Meetign of SCMR, Seattle, USA 2018
3 Beer M, Seyfarth T, Sandstede J, et al. Absolute concentrations of high-energy phosphate metabolites in normal, hypertrophied, and failing human myocardium measured noninvasively with 31 P-SLOOP magnetic resonance spectroscopy. Journal of the American College of Cardiology 2002;40:1267-74.
4 Neubauer S, Horn M, Cramer M, et al. Myocardial phosphocreatine-to-ATP ratio is a predictor of mortality in patients with dilated cardiomyopathy. Circulation 1997;96:2190-6.
5 Nascimben L, Ingwall JS, Lorell BH, et al. Mechanisms for increased glycolysis in the hypertrophied rat heart. Hypertension 2004;44:662-7.
6 Hettling H, van Beek JH. Analyzing the functional properties of the creatine kinase system with multiscale ‘sloppy’modeling. PLoS computational biology 2011;7:e1002130.
7 Wu F, Zhang EY, Zhang J, et al. Phosphate metabolite concentrations and ATP hydrolysis potential in normal and ischaemic hearts. The Journal of physiology 2008;586:4193-208.
8 Weiss RG, Gerstenblith G, Bottomley PA. ATP flux through creatine kinase in the normal, stressed, and failing human heart. Proceedings of the National Academy of Sciences of the United States of America 2005;102:808-13.
Figures