2306
Sex Differences in Stimulus Induced Blood Flow: The Importance of Sex Hormones1Université de Sherbrooke, Sherbrooke, QC, Canada, 2Bishop's University, Sherbrooke, QC, Canada
Synopsis
Sex differences in resting CBF has been reported, these differences may be explained through sex differences in sex hormones. There is currently no study that examines if this difference is maintained during stimulus-induced CBF. The current study evaluated men and naturally cycling women three times during their menstrual cycle at different sex hormone levels using a pCASL sequence. Preliminary results reveal sex differences in CBF response to the same stimulus, which is amplified when one considers sex hormones. These findings may reflect vascular effects of sex hormones, highlighting the importance of considering sex and hormone profiles when conducting fMRI
Introduction
Estrogen is a sex steroid that has the capacity to alter cebrovasculature: studies have almost consistently shown that women have higher resting cerebral blood flow (CBF) than men1–4, yet the reason behind this sex difference is unclear. In women, the menstrual cycle (MC) provides a natural fluctuation of estrogen not present in men and measuring CBF at different phases of their MC may provide insight on how different levels of estrogen can impact the brain's vascular system. Rat studies show that female rats have decreased vascular tone and respond less to stimulation compared to male rats5,6. These observations are attributed to estrogen’s ability to increase nitric oxide (NO) synthesis in vascular endothelial cells7–12. In fact, NO synthesis is higher in women than men and it fluctuates with estrogen levels13,14. Therefore, estrogen fluctuation could mediate stimulus-induced blood flow (SIBF). The objective of this study is to determine how the fluctuation of sex hormones during the MC influences SIBF. We hypothesize that men and women will experience similar SIBF response to a visual stimulus when women have low levels of estrogen and smaller SIBF when estrogen is high.Methods
We evaluated 4 healthy, normally cycling women at three phases in their MC (when estrogen and progesterone are low, when estrogen is high and progesterone is low, and when estrogen and progesterone are high) along with 5 healthy men. CBF was measured using a pCASL sequence (background suppression, label duration=1650ms, postlabel delay=1600ms, 2D mulcislice EPI readout, TR/TE 4000/14ms, 18 4mm, 3x3 resolution, FOV=240X240mm, effective temporal resolution 8000ms). Magnetic Resonance Angiography of the internal carotids and the vertebrate artery was used to improve placement of the labeling slab and increase consistency between repeated sessions. Visual stimulation (checkerboard on gray background) was presented in a 32 second on/off block design and regions of interest (ROI) were defined by correlating the raw CBF signal in each voxel with an HRF-convolved stimulus time-series and thresholding the map at r > 0.35. Hormonal assays of estradiol, progesterone were acquired at each testing session to classify each session.Results
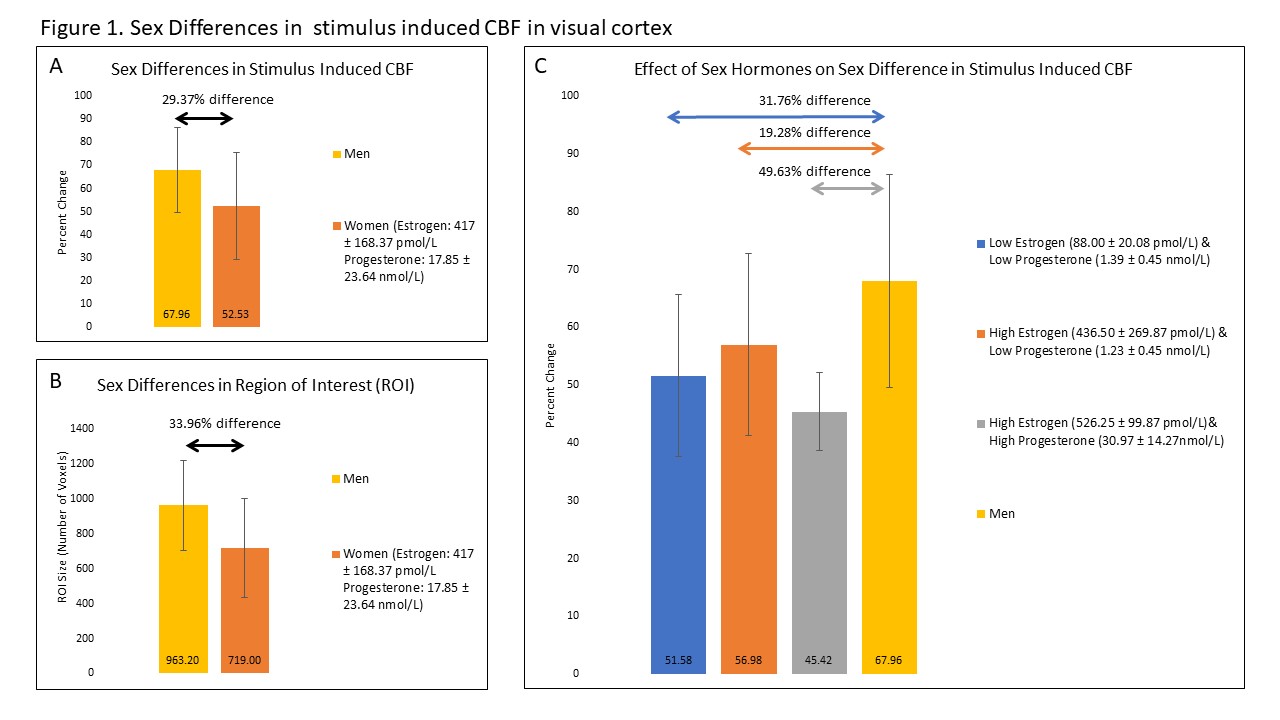
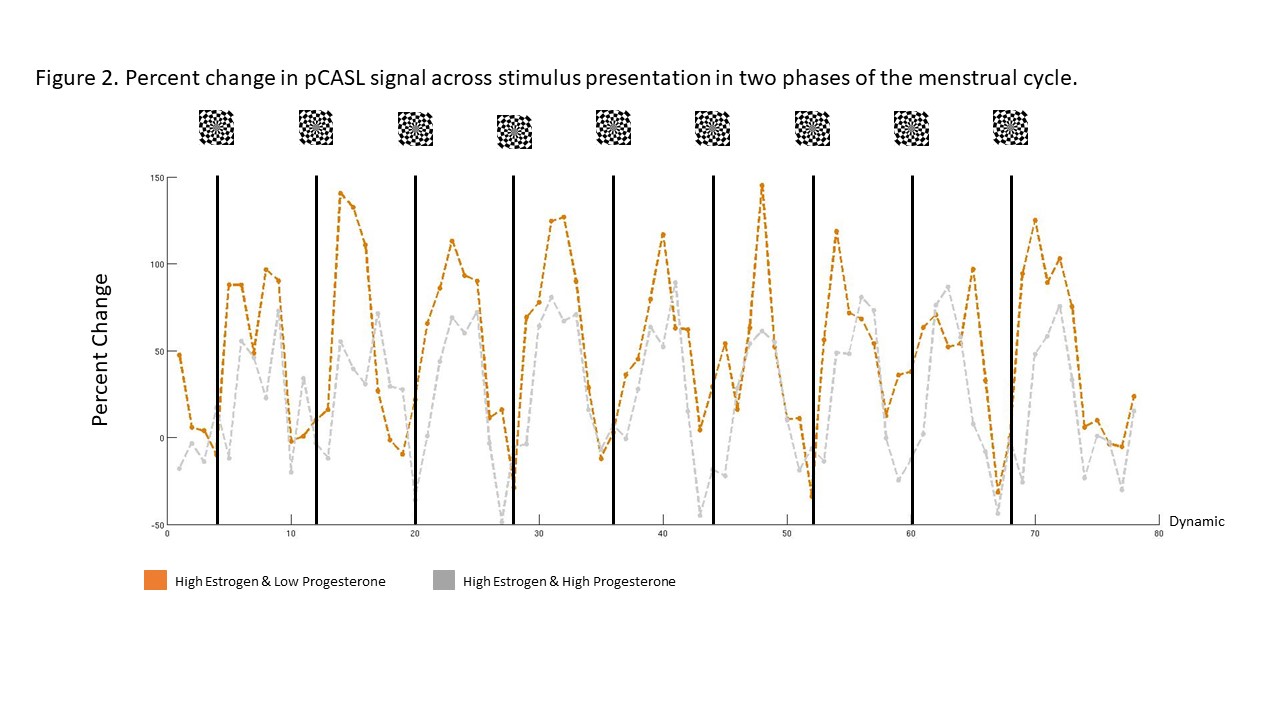
Without considering sex hormone level, women tended to display a smaller stimulus-evoked CBF (Figure 1a) and spatial activation extent (Figure 1b) compared to men. Here, CBF was 29.37% higher in men compared to women. When taking estrogen levels into account, this value reduced slightly to 19.28%, nevertheless, highlighting the suppressing effect of estrogen on stimulus-evoked CBF (Figure 1c). Unexpectedly, the effects of progesterone were far stronger than estrogen alone (49.63%), as illustrated in figure 2.Discussion
These preliminary data show that, overall, women show a smaller stimulus-evoked change in CBF as compared to men. This was expected given previous reports showing that estrogen is known to reduce vascular tone and reduced ability to dilate in response to stimulation5,15. This is likely a result of estrogen’s ability to increase NO production through chronic and acute exposure16. These results support that estrogen alters CBF response to stimulus in humans. In addition, we unexpectedly observed that SIBF was lowest when both estrogen and progesterone was high. The reason for this finding is unclear. One possibility is the result of a synergistic effect of both hormones on NO production. Meyer et al., (1993) showed that vascular tone is decreased and vascular response to a stimulus is reduced during late pregnancy in rats. This is a period in which both estrogen and progesterone levels are elevated17,18. Moreover, recent studies in human umbilical cord vascular endothelial cells demonstrated that combination doses of estrogen and progesterone enhance NO production by 1.6 fold compared to estrogen alone19,20. Alternatively, it is possible that the large decrease in cerebral reactivity when progesterone is high is due to progesterone’s neuronal effects. Progesterone’s metabolites have anxiolytic effects, reducing neuronal activity through GABAA receptor activity21–23. Hyperpolarization of the cell would reduce response to stimulation and may explain the decrease in SIBF.Conclusion
The result of the present study highlights the importance of considering sex and sex hormones while conducting hemodynamic-based measures of neural activity such as ASL and fMRI given that the latter relies on the interplay of neuronal activity and changes in blood flow, oxygenation, and volume24. Further investigation is required to understand the relationship between vascular structure, reactivity and sex hormones. Additional investigations to evaluate resting CBF, arterial diameters and stimulus-induced dilation in arteries across the MC and between the sexes is currently being conducted. These measures will help clarify the role of cerebral vascular structure on differences in SIBF in sex differences.Acknowledgements
No acknowledgement found.References
1. Kastrup A, Li T-Q, Glover GH, Kruger G, Moseley ME. Gender Differences in Cerebral Blood Flow and Oxygenation Response During Focal Physiologic Neural Activity. J Cereb Blood Flow Metaholism. 19:1066-1. http://journals.sagepub.com/doi/pdf/10.1097/00004647-199910000-00002. Accessed November 6, 2017.
2. Gur RE, Gur RC. Gender differences in regional cerebral blood flow. Schizophr Bull. 1990;16(2):247-254. doi:10.1093/schbul/16.2.247.
3. Rodriguez G, Warkentin S, Risberg J, Rosadini G. Sex differences in regional cerebral blood flow. J Cereb Blood Flow Metab. 1988;8(6):783-789. doi:10.1038/jcbfm.1988.133.
4. Parkes LM, Rashid W, Chard DT, Tofts PS. Normal Cerebral Perfusion Measurements Using Arterial Spin Labeling: Reproducibility, Stability, and Age and Gender Effects. Magn Reson Med. 2004;51(4):736-743. doi:10.1002/mrm.20023.
5. Geary GG, Krause DN, Duckles SP. Estrogen reduces myogenic tone through a nitric oxide-dependent mechanism in rat cerebral arteries. Am J Physiol. 1998;275(1 Pt 2):H292-300. http://www.ncbi.nlm.nih.gov/pubmed/9688926.
6. Geary GG, Krause DN, Duckles SP. Estrogen reduces mouse cerebral artery tone through endothelial NOS- and cyclooxygenase-dependent mechanisms. Am J Physiol Heart Circ Physiol. 2000;279(2):H511-9. http://www.ncbi.nlm.nih.gov/pubmed/10924048.
7. Duckles SP, Krause DN. Cerebrovascular effects of oestrogen: Multiplicity of action. Clin Exp Pharmacol Physiol. 2007;34(8):801-808. doi:10.1111/j.1440-1681.2007.04683.x.
8. Krause DN, Duckles SP, Pelligrino D a. Influence of sex steroid hormones on cerebrovascular function. J Appl Physiol. 2006;101(4):1252-1261. doi:10.1152/japplphysiol.01095.2005.
9. Miller VM, Duckles SP. Vascular actions of estrogens: functional implications. Pharmacol Rev. 2008;60(2):210-241.
10. Ospina J a, Duckles SP, Krause DN. 17beta-estradiol decreases vascular tone in cerebral arteries by shifting COX-dependent vasoconstriction to vasodilation. Am J Physiol Heart Circ Physiol. 2003;285(1):H241-H250. doi:10.1152/ajpheart.00018.2003.
11. Pelligrino D a, Galea E. Estrogen and cerebrovascular physiology and pathophysiology. Jpn J Pharmacol. 2001;86(2):137-158. doi:10.1254/jjp.86.137.
12. Toda N, Ayajiki K, Okamura T. Cerebral blood flow regulation by nitric oxide : Pharmacol Rev. 2009;61(1):62-97. doi:10.1124/pr.108.000547.62.
13. Rosselli M, Imthurm B, Macas E, Keller PJ, Dubey RK. Circulating nitrite/nitrate levels increase with follicular development: indirect evidence for estradiol mediated NO release. Biochem Biophys Res Commun. 1994;202(3):1543-1552. doi:10.1006/bbrc.1994.2107.
14. Forte P, Kneale BJ, Milne E, et al. Evidence for a Difference in Nitric Oxide Biosynthesis Between Healthy Women and Men. 1998:730-734.
15. Meyer MC, Brayden JE, McLaughlin MK. Characteristics of vascular smooth muscle in the maternal resistance circulation during pregnancy in the rat. Am J Obstet Gynecol. 1993;169(6):1510-1516. doi:10.1016/0002-9378(93)90427-K.
16. Stirone C, Boroujerdi A, Duckles SP, Krause DN. Estrogen receptor activation of phosphoinositide-3 kinase, akt, and nitric oxide signaling in cerebral blood vessels: rapid and long-term effects. Mol Pharmacol. 2005;67(1):105-113. doi:10.1124/mol.104.004465.receptor.
17. Hashimoto I, Henricks DM, Anderson LL, Melampy RM. Progesterone and pregn-4-en-20 alpha-ol-3-one in ovarian venous blood during various reproductive states in the rat. Endocrinology. 1968;82(2):333-341.
18. Yoshinaga K, Hawkins RA, Stocker JF. Estrogen secretion by the rat ovary in vivo during the estrous cycle and pregnancy. Endocrinology. 1969;85(1):103-112. doi:10.1210/endo-85-1-103.
19. Pang Y, Thomas P. Additive effects of low concentrations of estradiol-17β and progesterone on nitric oxide production by human vascular endothelial cells through shared signaling pathways. J Steroid Biochem Mol Biol. 2017;165:258-267. doi:10.1016/j.jsbmb.2016.06.014.
20. Thomas P, Pang Y. Protective actions of progesterone in the cardiovascular system: Potential role of membrane progesterone receptors (mPRs) in mediating rapid effects. Steroids. 2013;78(6):583-588. doi:10.1016/j.steroids.2013.01.003.
21. Stell BM, Brickley SG, Tang CY, Farrant M, Mody I. Neuroactive steroids reduce neuronal excitability by selectively enhancing tonic inhibition mediated by delta subunit-containing GABAA receptors. Proc Natl Acad Sci U S A. 2003;100(24):14439-14444. doi:10.1073/pnas.2435457100.
22. Smith SS. Progesterone administration attenuates excitatory amino acid responses of cerebellar Purkinje cells. Neuroscience. 1991;42(2):309-320. doi:10.1016/0306-4522(91)90377-Z.
23. Bitran D, Purdy RH, Kellog CK. Anxiolytic effect of progesterone is associated with increases in cortical alloprenanolone and GABAA receptor function. Pharmacol Biochem Behav. 1993;45(2):423-428. doi:10.1016/0091-3057(93)90260-Z.
24. Logothetis NK, Wandell B a. Interpreting the BOLD signal. Annu Rev Physiol. 2004;66:735-769. doi:10.1146/annurev.physiol.66.082602.092845.
Figures