2228
Dependence of rNOE-CEST signals on molecular weight1Division of Medical Physics in Radiology, German Cancer Research Center (DKFZ), Heidelberg, Germany, 2Molecular Structure Analysis, German Cancer Research Center (DKFZ), Heidelberg, Germany
Synopsis
In this study, rNOE-CEST signals of proteins have been demonstrated to depend on molecular weight. This finding can explain the observed intensity decrease of aliphatic rNOE-CEST signals in tumors in comparison to healthy tissue.
Introduction
So-called relayed nuclear Overhauser effect (rNOE) chemical exchange saturation transfer (CEST) signals have been shown to provide information about cancer tissue, e.g. tumor grading1 or the differentiation between radiation necrosis and tumor progression2. In brain tumors, a significant decrease of rNOE-CEST signals in comparison to healthy tissue has been consistently reported by several groups1-6. However, the underlying origin of the observed signal decrease remains to be clarified. rNOE-CEST signals are generally assumed to originate from bulk mobile macromolecules, in particular proteins and phospholipids. The signal decrease in tumors cannot simply be attributed to a reduction in the concentration of mobile macromolecules since the proportion of mobile proteins/peptides is upregulated in cancer tissue7. Recently, a correlation of rNOE-CEST signals with the folding state of proteins has been reported, in principle, providing an explanation for the observed signal decrease8-10. In this study, we complement our previous studies on the physical mechanisms of rNOE-CEST signals with an investigation of the dependence of rNOE-CEST signals on molecular weight (MW).Methods
Experiments were performed on a 14.1 T NMR spectrometer (Bruker, Germany) employing a continuous wave (cw) pre-saturation of duration 17 s and amplitude B1 = 1 µT. The duration of pre-saturation complied with the criterion for steady-state (ss) measurements. The temperature of the samples was stabilized at 37 °C using the internal heating and cooling device.
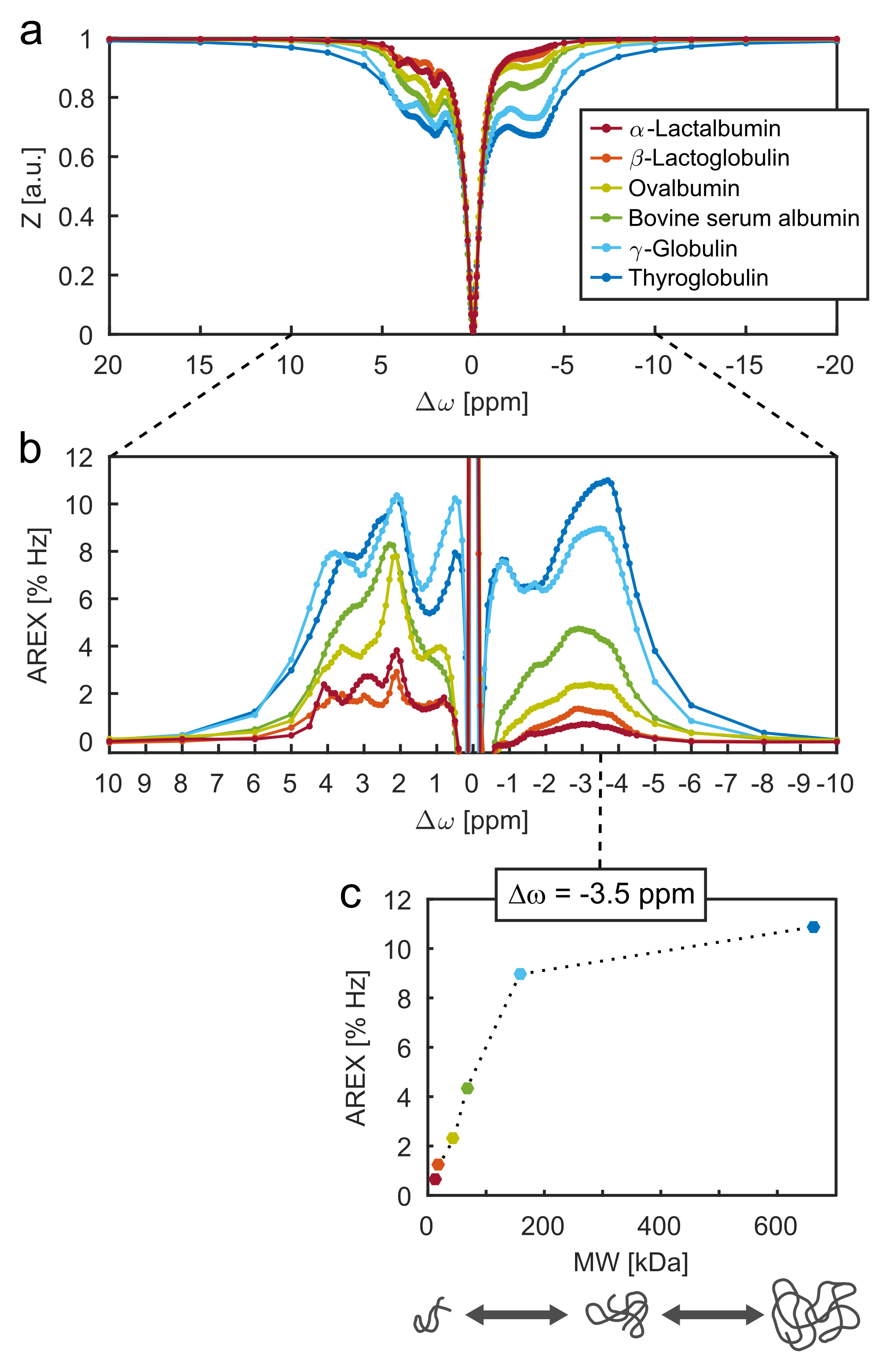
Six model solutions were prepared containing mobile globular proteins of different MWs in the range of 14 to 660 kDa. The individual protein types are listed in the legend of Fig. 1a. Protein content was kept constant at a proportion of 10% (w/v). This means that the molar concentration of proteins was lower for heavier molecular weights. Model solutions were buffered at pH 7.1 using phosphate-buffered saline (PBS).
Isolated CEST signals were calculated by the apparent exchange-dependent relaxation rate11, AREX = 1/T1 ∙ (1/Z – 1/Zref), employing a bi-Lorentzian fit of the direct water saturation and the semi-solid magnetization transfer (ssMT) as the reference value Zref.
Results
The rNOE-CEST signals of aliphatic protons in proteins show a distinct dependence on MW (Fig. 1c). At MWs less than ca. 100 kDa, a linear increase of the signal strength as a function of MW was observed. Above ca. 100 kDa, the increase diminishes and the signal strength levels off. Also around 100 kDa, a broad signal with a linewidth comparable to ssMT effects starts to show up (Fig. 1a, blue lines, Δω ≥ 8 ppm and ‒8 ppm ≥ Δω).Discussion
A dependence of rNOE-CEST signals on MW is in line with expectations. The process of rNOE-CEST involves an intramolecular magnetization transfer from non-labile (e.g. CHx) to chemically exchangeable protons (e.g. NHx, OH)12. This intramolecular magnetization transfer is mediated by a cascade of dipolar couplings and is referred to as spin diffusion13. As dipolar couplings are known to strongly depend on the tumbling of molecules (i.e. molecular correlation time), a dependence of rNOE-CEST signals on MW can be expected.
The diminution of the increase of rNOE-CEST signals above ca. MW > 100 kDa is most likely caused by decreasing transversal relaxation times (T2). Of course, the actual magnetization transfer is more effective at higher MWs but the observable signal strength cannot increase by the same extent due to shorter T2 and hence line broadening10. This agrees with the observed rise of a broad ssMT-like effect for MW > 100 kDa. At MW > 660 kDa (not explored in this study), we believe that selective rNOE-CEST signals are almost completely transformed into a single broad ssMT resonance.
Effects from different folding states of the proteins are negligible in this study because all of the investigated proteins belong to the same class of globular proteins which are spherical in shape.
Conclusion
The dependence of rNOE-CEST signals on MW was verified experimentally by the investigation of different globular proteins. It was demonstrated that a decrease of the rNOE-CEST signals at constant protein content can be caused by reduced MWs. Accordingly, the observed drop of aliphatic rNOE-CEST signals in tumors can be explained by an increased portion of mobile proteins/peptides which are comparatively small in size (MW < 100 kDa).Acknowledgements
No acknowledgement found.References
1. Jones CK, Huang A, Xu J, et al. Nuclear Overhauser enhancement (NOE) imaging in the human brain at 7 T. Neuroimage. 2013;77:114-124.
2. Mehrabian H, Desmond KL, Soliman H, et al. Differentiation between Radiation Necrosis and Tumor Progression Using Chemical Exchange Saturation Transfer. Clin Cancer Res. 2017; 23(14):3667-3675.
3. Xu J, Zaiss M, Zu Z, et al. On the origins of chemical exchange saturation transfer (CEST) contrast in tumors at 9.4 T. NMR Biomed. 2014; 27(4):406-416.
4. Zaiss M, Windschuh J, Paech D, et al. Relaxation-compensated CEST-MRI of the human brain at 7 T: Unbiased insight into NOE and amide signal changes in human glioblastoma. Neuroimage. 2015; 112:180-188.
5. Heo H-Y, Jones CK, Hua J, et al. Whole-Brain Amide Proton Transfer (APT) and Nuclear Overhauser Enhancement (NOE) Imaging in Glioma Patients Using Low-Power Steady-State Pulsed Chemical Exchange Saturation Transfer (CEST) Imaging at 7T. J Magn Reson Imaging. 2016;44:41-50.
6. Desmond KL, Mehrabian H, Chavez S, et al. Chemical Exchange Saturation Transfer for Predicting Response to Stereotactic Radiosurgery in Human Brain Metastasis. Magn Reson Med. 2017; 78(3):1110-1120.
7. Yan K, Fu Z, Yang C, et al. Assessing Amide Proton Transfer (APT) MRI Contrast Origins in 9 L Gliosarcoma in the Rat Brain Using Proteomic Analysis. Mol Imaging Biol. 2015; 17(4):497-487.
8. Zaiss M, Kunz P, Goerke S, et al. MR imaging of protein folding in vitro employing Nuclear-Overhauser-mediated saturation transfer. NMR Biomed. 2013; 26(12):1815-1822.
9. Goerke S, Zaiss M, Kunz P, et al. Signature of protein unfolding in chemical exchange saturation transfer imaging. NMR Biomed. 2015; 28(7):906-913
10. Goerke S, Milde K, Zaiss M, et al. Aggregation-induced changes in the chemical exchange saturation transfer (CEST) signals of proteins. NMR Biomed. 2016; 30(1):e3665.
11. Zaiss M, Xu J, Goerke S, et al. Inverse Z-spectrum analysis for spillover-, MT-, and T1-corrected steady-state pulsed CEST-MRI – application to pH-weighted MRI of acute stroke. NMR Biomed. 2014;27(3):240-252.
12. van Zijl PCM, Zhou J, Mori N, et al. Mechanism of Magnetization Transfer During On-Resonance Water Saturation. A New Approach to Detect Mobile Proteins, Peptides, and Lipids. Magn Reson Med. 2003;49:440-449.
13. Neuhaus D, and Williamson MP. The Nuclear Overhauser Effect in Structural and Conformational Analysis. Wiley. 1989.
Figures