0532
Early specific connections of the Visual Word Form Area during the first year of reading instruction: a longitudinal MRI study in children1INSERM, Gif-sur-Yvette, France, 2INSERM, Paris, France, 3CEA, Gif-sur-Yvette, France, 4University College London, London, United Kingdom, 5College de France, Paris, France, 6CNRS, Gif-sur-Yvette, France
Synopsis
Specialized in word recognition, the visual word form area (VWFA) is located in the posterior fusiform gyrus of the left hemisphere, regardless of one’s writing system. To test the hypothesis that this consistent location is determined by specific connections to linguistic regions, we studied 6-7year old children throughout the process of learning to read. Using diffusion and functional MRI, we analysed the white-matter connectivity of cortical regions processing visual categories (words, houses, faces, tools). We showed that the emerging VWFA has specific and stable connections to the inferior-dorsal parietal cortex, and these connections exhibit microstructural maturation related to reading improvement.
Introduction
Specialized in word recognition, the visual word form area (VWFA) is a region systematically located in the ventral occipito-temporal cortex of the left hemisphere1, within the posterior fusiform gyrus, regardless of one’s language and writing system2. The consistent location of the VWFA3 in the mosaic of areas processing visual categories (e.g. houses, faces, tools) may be due to specific long-range connections to other regions of the language network, which are absent for the surrounding cortex4. However, only one study has investigated this hypothesis in children through the cortical terminations of VWFA connections5. Here we aimed to explore the entirety of white-matter connectivity of category-specific regions in terms of specificity (i.e. word compared to house, face and tool areas) and stability throughout the process of learning to read. Moreover, we assessed how the microstructural properties of these specific connections evolve with reading improvement.Methods
Ten children (5 females) were recruited in a longitudinal study. Participants were evaluated behaviourally, and underwent MRI scanning (3T Siemens TRIO) at six different time points spanning the beginning and end of the first school year of reading instruction. Diffusion MRI data (45 directions, b=1000s/mm2), which constitute the focus of this study, were obtained at the first and last time points (mean ages: 6.2±0.3 years, 7.2±0.4 years), while reading performance (number of words read in one minute) and fMRI data were obtained at each time point.
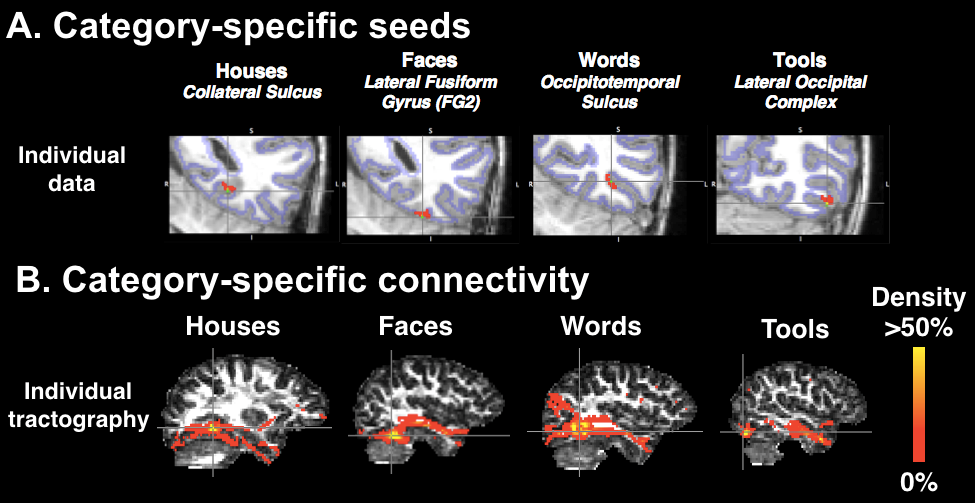
All functional data were used to localize functional peaks corresponding to the already developed house, face, and tool processing areas along with the emerging VWFA over the first year of reading instruction. Functional peaks were used to generate tractography seeds for the first and last time points (Figure 1A). To tentatively resolve simple crossing fiber configurations, DW images were analyzed based on a fourth order analytical Q-ball model6. Individual whole-brain 3D tractography was performed with an algorithm using regularized particle trajectories7. Tracts were converted into density maps (Figure 1B), normalized onto a population template constructed from diffusion tensor images using DTI-TK8, and smoothed with a 3mm Gaussian Kernel. For each visual category, these subject-level density maps were binarized and averaged over subjects to create a group-level probability map.
To test the stability and specificity of the connectivity for each visual category processing area, we performed a voxel-wise ANCOVA (FSL’s randomise9) on density maps with two within-subject factors (time point and visual category), and two within-subject covariates (age and reading score). Finally, we tested the hypothesis that only specific connections for the word processing region shows microstructural changes related to improvement in reading performances. The specific connectivity of each category (resulting from previous ANCOVA) was then used as a region-of-interest to extract average fractional anisotropy (FA), axial diffusivity (AD), and radial diffusivity (RD) weighted by the group-level probability map. We computed partial correlations between changes in each DTI parameter and changes in reading score from the beginning to the end of the school year, while controlling for the age increase.
Results
Connections emanating from the emerging VWFA in children were stable across time points, suggesting that they antedated reading instruction. Compared with other visual categories, these tracts had preferential long-distance connectivity to the left inferior, dorsal parietal cortex (Figure 2A). Most importantly, only these connections exhibited significant maturational changes in line with reading improvements through decreasing RD (Figure 2B).Discussion
The parietal cortex is known to intervene in healthy adults when they read words in unfamiliar and novel formats, such as when distorted through rotation, increased letter spacing, hemifield shifting, or low contrast10. Behaviorally, this is translated by an effortful, not automated reading. Children’s reliance on letter-by-letter reading, a stepping stone in literacy, may thus explain the location of the VWFA through its connectivity to the parietal cortex at this early stage11. This is further corroborated by the fact that only in the VWFA connectivity did improvements in reading correlate with a decrease in RD, a parameter reflecting myelination. In our child cohort, it thus seems that the specific connections between the word-specific region and the parietal cortex are strongly involved in the learning process and mediate crucial information during the first stages of learning to read, which leads to their specific myelination.Conclusion
These findings support children’s transient reliance on the parietal lobe for attentional control over letter-by-letter reading8 and disambiguation of mirrors letters (e.g. b vs d). The role of these connections is likely maximal at the onset of learning to read when parietal resources are highly solicited to manage the strong attentional demands, needed to decipher words.Acknowledgements
This research was supported by grants from the Fondation Bettencourt-Schueller and the Fondation de France.References
1. Dehaene, et al. Illiterate to literate: behavioural and cerebral changes induced by reading acquisition. Nat. Rev. Neurosci. 2015;16:234–44.
2. Rueckl et al. Universal brain signature of proficient reading: Evidence from four contrasting languages. PNAS: 2015; 112, 15510-115515
3. Jobard, et al. Evaluation of the dual route theory of reading: A metanalysis of 35 neuroimaging studies. Neuroimage: 2003;20, 693–712.
4. Hannagan, et al. Origins of the specialization for letters and numbers in ventral occipitotemporal cortex. Trends Cogn. Sci. 2015;19:374–382.
5. Saygin, et al. Connectivity precedes function in the development of the visual word form area. Nat. Neurosci. 2016:19:1250–5.
6. Descoteaux et al. Regularized, fast, and robust analytical Q-ball imaging. Magn. Reson. Med: 2007:58, 497–510.
7. Perrin, M et al. Fiber tracking in q-ball fields using regularized particle trajectories. Inf. Process. Med. Imaging 2005:19;52–63.
8. Zhang, et al. Deformable registration of diffusion tensor MR images with explicit orientation optimization. Med. Image Anal. 2006:10;764–85.
9. Winkler, et al Permutation inference for the general linear model. NeuroImage, 2014;92:381-397.
10. Cohen, et al. Reading normal and degraded words: Contribution of the dorsal and ventral visual pathways. Neuroimage 2008:40;353–366.
11. Dehaene-Lambertz, G., Dehaene, S. The emergence of the visual word form area: Longitudinal evolution of category-specific responses in ventral visual cortex during reading acquisition. Submitted.
Figures