0151
High-frequency BOLD responses in human thalamus detected through fast fMRI at 7 Tesla1Athinoula A. Martinos Center for Biomedical Imaging, Massachusetts General Hospital, Boston, MA, United States, 2Society of Fellows, Harvard University, Cambridge, MA, United States, 3Radiology, Harvard Medical School, boston, MA, United States, 4Radiology, Harvard Medical School, Boston, MA, United States
Synopsis
No current technique can noninvasively localize neural activity in human subcortical structures at subsecond temporal resolution. Recent studies have demonstrated that fast (>0.2 Hz) fMRI responses can be detected in human cortex. We aimed to test whether fast fMRI signals can also be detected in the thalamus. We presented oscillating visual stimuli in order to induce oscillatory neural activity in visual thalamus, and observed large-amplitude fMRI oscillations at 0.5 Hz. We conclude that high-frequency fMRI responses can be detected in thalamus, suggesting fast fMRI has the potential to be used for whole-brain imaging.
Introduction
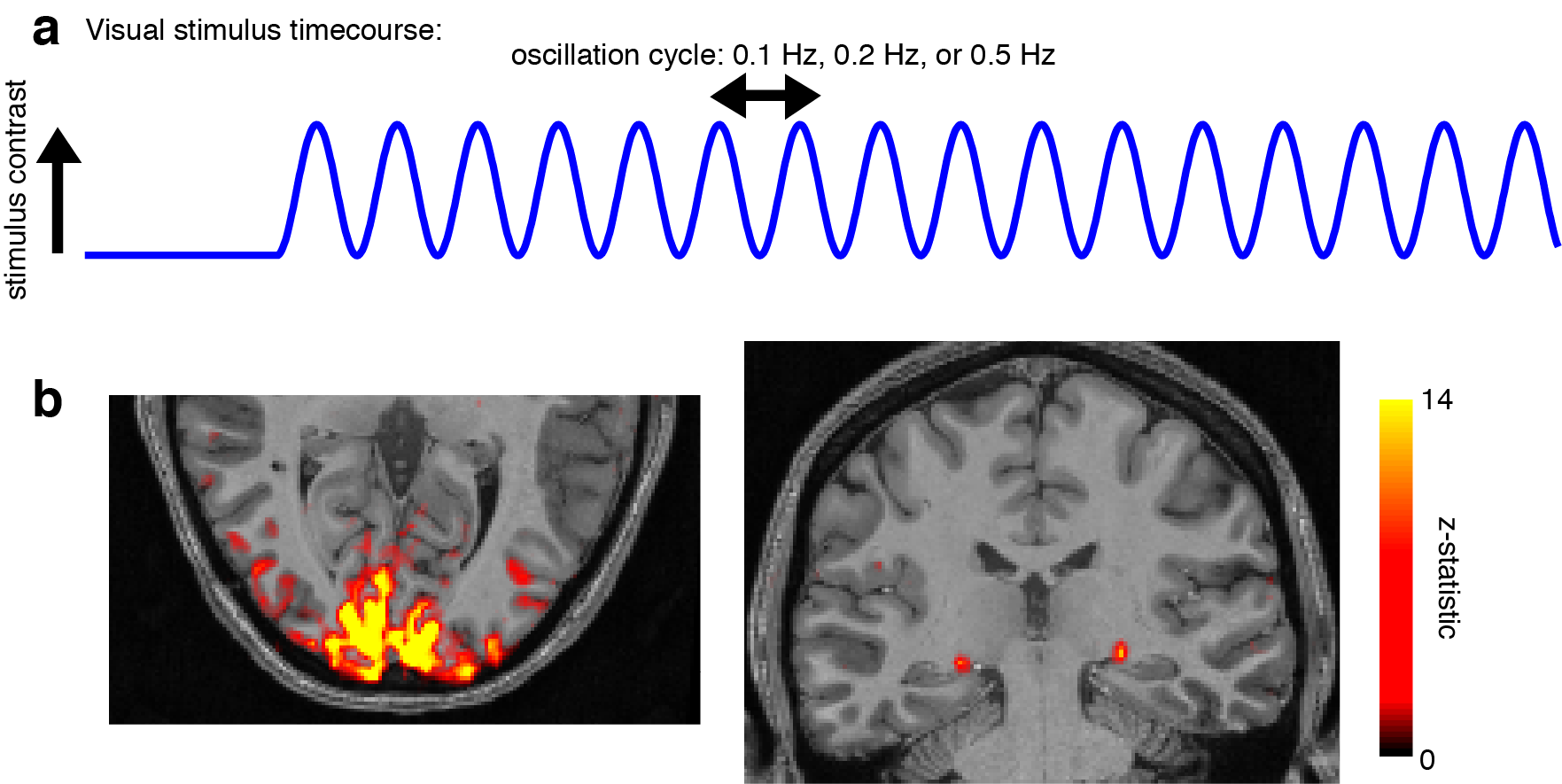
A key strength of fMRI is its ability to image the whole brain, including subcortical structures such as thalamus that are challenging to access through other techniques. This imaging is typically performed at low sampling rates, as the hemodynamic response is classically assumed to be slow, but recent studies have demonstrated that surprisingly large >0.2 Hz signals can be detected in human cortex, both in task-driven [1] and resting state [2-6] contexts. Subcortical fMRI responses are more challenging to measure, since the small size of subcortical nuclei and the low signal-to-noise in the center of the brain reduces detectability. Furthermore, the shape and timing of the hemodynamic response varies substantially over the brain [7-10], and whether fast hemodynamics can occur in deep structures is not known. We aimed to test whether high-frequency fMRI signals can be detected in human thalamus. We studied this using oscillatory visual stimuli to drive oscillating neural activity in primary visual cortex (V1) and the lateral geniculate nucleus (LGN, a visual nucleus of the thalamus).Methods
Five subjects gave informed consent and were scanned on a 7T Siemens scanner with a custom-built 32-channel head coil array. Each session began with a 0.75 mm isotropic multi-echo MPRAGE [11] and initial locations for V1 and LGN were identified using anatomical landmarks. Functional runs consisted of 15 oblique slices that were positioned to include both LGN and V1, acquired as single-shot gradient-echo blipped-CAIPI SMS-EPI [12] with 2 mm isotropic resolution (R=2 acceleration, MultiBand factor=3, matrix=120×120, CAIPI shift=FOV/3, TR=227 ms, TE=24 ms, echo-spacing=0.59 ms, flip angle=30°). Visual stimuli were presented for four continuous minutes, in which a flickering radial checkerboard inverted at 12 Hz. The luminance contrast of the checkerboard oscillated at either 0.2 Hz or 0.5 Hz in each run (Fig. 1a). Subjects performed a simple detection task to maintain fixation throughout the run. For analysis, regions of interest (ROIs) in V1 and LGN were defined using functional maps from an initial localizer run using 0.1 Hz stimulation (Fig. 1b), with an anatomical constraint. The frequency response (FR) was calculated as the ratio of the amplitude of the 0.5 Hz response to the 0.2 Hz response, times 100. Oscillation amplitude was calculated as the magnitude of the best-fit sine wave. 95% confidence intervals (CI) for the oscillation amplitude were calculated via bootstrap, resampling subjects and cycles 1000 times.Results
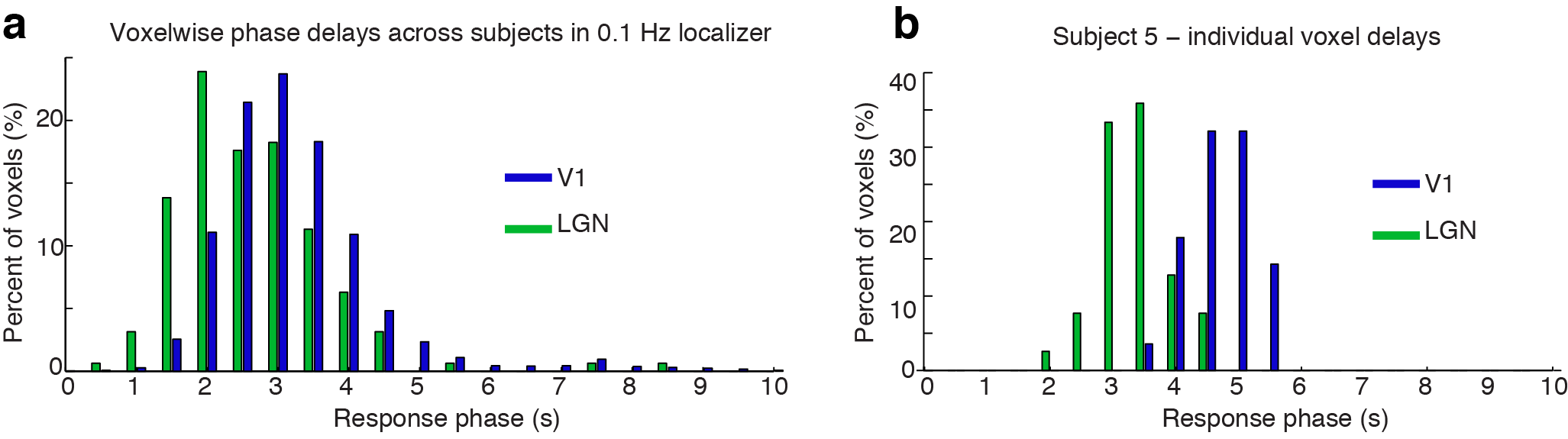
We first analyzed mean fMRI responses to the 0.2 Hz stimulus, and observed robust fMRI oscillations both in V1 and LGN (Fig. 2a, V1 amplitude=1.26%, CI=[0.95 1.61]; LGN amplitude: 1.04%, CI=[0.71 1.38]). Canonical models of the hemodynamic response would then predict undetectable responses at 0.5 Hz, with a relative frequency response (FR) of 1.3, which would lead to oscillations with magnitude <0.02%. However, consistent with prior studies [1], we observed oscillations at 0.5 Hz in V1 that were substantially larger than the canonical prediction (Fig. 2b, V1 amplitude=0.11%, CI=[0.07 0.16]). In addition, 0.5 Hz oscillations of similar magnitude were observed in LGN (Fig. 2b, LGN amplitude=0.07%, CI=[0.01 0.13]). These results corresponded to an FR of 6.6 in LGN and 9.4 in V1, several times larger than the 1.3 FR of canonical models. We further observed that the response in LGN consistently preceded that of V1 by hundreds of milliseconds (peak 0.2 Hz timing: 1.18 s in V1, 0.82 s in LGN; 0.5 Hz timing: 1.48 s in V1, 1.10 s in LGN). Plotting voxel-level temporal delays demonstrated that the population of LGN voxels typically responded earlier than V1 by hundreds of milliseconds (Fig. 3a). In some individual subjects, these temporal delay distributions were non-overlapping across V1 and LGN (Fig. 3b), suggesting that hemodynamic responses consistently occur with an earlier onset in LGN than in V1.Discussion
Our results demonstrate that >0.2 Hz fMRI signals are present and robustly detected in human visual thalamus (LGN). The relative frequency response was very similar across V1 and LGN, suggesting similar hemodynamic nonlinearities exist in both: specifically, fMRI responses to high-frequency stimuli are larger and faster than predicted by responses to conventional block-design stimuli. However, the response phase was consistently different across regions, with LGN oscillations peaking hundreds of milliseconds earlier than V1. This fMRI response delay is much larger than the expected neural delay (as V1 neural activity lags LGN by only tens of milliseconds [13]) and is therefore likely due to faster hemodynamics in LGN. We conclude that fast fMRI can detect high-frequency signals in human thalamus, and suggest that mapping local hemodynamic delays may be crucial for analyzing whole-brain fast fMRI data.Acknowledgements
This work was funded by NIH grants K99-MH111748, R01-EB019437, and P41-EB015896; and NCRR shared resource instrumentation grants S10-RR023401, S10-RR023403, and S10-RR020948.References
1. Lewis LD, Setsompop K, Rosen BR, Polimeni JR (2016) Fast fMRI can detect oscillatory neural activity in humans. Proc Natl Acad Sci USA 113:E6679–E6685. doi: 10.1073/pnas.1608117113
2. Lee H-L, Zahneisen B, Hugger T, et al (2013) Tracking dynamic resting-state networks at higher frequencies using MR-encephalography. Neuroimage 65:216–222. doi: 10.1016/j.neuroimage.2012.10.015
3. Chen JE, Glover GH (2015) BOLD fractional contribution to resting-state functional connectivity above 0.1 Hz. Neuroimage 107:207–218. doi: 10.1016/j.neuroimage.2014.12.012
4. Trapp C, Vakamudi K, Posse S (2017) On the detection of high frequency correlations in resting state fMRI. Neuroimage 1–12. doi: 10.1016/j.neuroimage.2017.01.059
5. Kalcher K, Boubela RN, Huf W, et al (2014) The Spectral Diversity of Resting-State Fluctuations in the Human Brain. PLoS ONE 9:e93375. doi: 10.1371/journal.pone.0093375.t003
6. Gohel SR, Biswal BB (2015) Functional integration between brain regions at rest occurs in multiple-frequency bands. Brain Connectivity 5:23–34. doi: 10.1089/brain.2013.0210
7. Miezin FM, Maccotta L, Ollinger JM, et al (2000) Characterizing the Hemodynamic Response: Effects of Presentation Rate, Sampling Procedure, and the Possibility of Ordering Brain Activity Based on Relative Timing. Neuroimage 11:735–759. doi: 10.1006/nimg.2000.0568
8. Handwerker DA, Ollinger JM, D'Esposito M (2004) Variation of BOLD hemodynamic responses across subjects and brain regions and their effects on statistical analyses. Neuroimage 21:1639–1651. doi: 10.1016/j.neuroimage.2003.11.029
9. Saad ZS, Ropella KM, Cox RW, DeYoe EA (2001) Analysis and use of FMRI response delays. Human brain mapping 13:74–93.
10. de Zwart JA, Silva AC, van Gelderen P, et al (2005) Temporal dynamics of the BOLD fMRI impulse response. Neuroimage 24:667–677. doi: 10.1016/j.neuroimage.2004.09.013
11. van der Kouwe AJW, Benner T, Salat DH, Fischl B (2008) Brain morphometry with multiecho MPRAGE. Neuroimage 40:559–569. doi: 10.1016/j.neuroimage.2007.12.025
12. Setsompop K, Gagoski BA, Polimeni JR, et al (2012) Blipped-controlled aliasing in parallel imaging for simultaneous multislice echo planar imaging with reduced g-factor penalty. Magn Reson Med 67:1210–1224. doi: 10.1002/mrm.23097
13. Rockland KS, Kaas JH, Peters A (1997) Extrastriate cortex in primates. Plenum Press
Figures