5370
Physiological noise correction improves sensitivity, specificity, and reproducibility of resting-state functional connectivity in a reading model1Dept. of Neurology, Emory University, Atlanta, GA, United States, 2Center for Visual and Neurocognitive Rehabilitation, Atlanta VAMC, Decatur, GA, United States, 3Center for Advanced Brain Imaging, GSU/GT, Atlanta, GA, United States, 4Dept. of Physics & Astronomy, Georgia State University, Atlanta, GA, United States, 5Dept. of Educational Psychology, Special Education, and Communication Disorders, Georgia State University, Atlanta, GA, United States, 6Dept. of Psychology, Georgia State University, Atlanta, GA, United States
Synopsis
Amongst several sources of noise, physiological noise (PN) from cardiac and respiratory cycles affects reliable quantification of rsFC measures such as correlation coefficient (CC). The purpose of this study is to determine the effects of PN on specificity, sensitivity and reproducibility of rsFC maps in a ‘reading’ model. We show that a combination of multiple methodologies to correct for such noise leads to improved signal fluctuations (tSNR) that culminates in higher specificity and sensitivity to neuronal fluctuations that are closer to actual ground truth. Applying our methodologies to a ‘reading’ model, we show that, irrespective of session, correction for PN results in meaningful discrimination of reading networks between typical and struggling readers.
Purpose
Resting-state functional connectivity (rsFC) MRI has emerged as a powerful tool to investigate diseased populations as it can be agnostic to task bias, does not require subjects to perform a task, and is simple to acquire. However, its practical use in the clinical setting requires improving reliability and reproducibility of the rsFC signals. Amongst several sources of noise1, physiological noise (PN) from cardiac and respiratory cycles affects reliable quantification of rsFC measures such as correlation coefficient (CC)2,3. The purpose of this study is to determine the effects of PN on specificity, sensitivity and reproducibility of rsFC maps in a ‘reading’ model.Methods
Subjects: Eight typical and eight struggling adult readers were recruited from the Center for the Study of Adult Literacy, and classified based on reading assessments. MRI: High-resolution T1-weighted MPRAGE and rsfMRI (TR=2sec, TE=30ms, voxel=3.4x3.4x4mm3, 32 slices) images were acquired on a Siemens 3T Tim Trio with a 12 ch-head coil during two sessions spaced 2-4 weeks apart. Pulse oximetry was used to quantify the mean heart beat per minute (MBPM), and pulmonary plethysmography was used to quantify respiratory volume per time (RVT)4. Pre-processing of rsfCMRI: The rsFC images were corrected for slice timing, global head motion, EPI distortions, and spatially normalized to MNI, followed by masking of the ventricles, low-pass filtering between 0.001 and 0.1Hz, and spatially smoothed (FWHM=6mm). Since global signal regression (GSR) has been shown to artificially center the CC distribution around zero5, we did not include GSR. We incorporated RETROICOR6 to minimize global PN, and applied ANATICOR7 to minimize local white matter BOLD fluctuations. The RVT and MBPM signals were voxel-wise detrended to minimize BOLD-like vasomotor nuisance signals. The PN correction was pre-processed eight different ways: no PN correction (No Physio), RETROICOR-only, ANATICOR-only, RETROICOR+ANATICOR, RVTMBPM-only, RETROICOR+RVTMBPM, RVTMBPM+ANATICOR, and RETROICOR+RVTMBPM+ANATICOR. Post-processing of rsfCMRI: Seed-based CC analysis was applied in a whole-brain manner, and transformed using a Fisher Z-transform. For each of the eight combinations, we quantified seed temporal Signal to Noise Ratio (tSNR), sensitivity to apriori expected connection (e.g. L-STG to R-STG) via modeling of the rsFC CC with tSNR, specificity of expected rsFC map using dice coefficient (DC)8, and Intra-class correlation coefficient (ICC)9 to assess reproducibility. Finally, we compare the reproducible networks between average and poor readers. The CC analysis was also carried out for right hemisphere (RH) seeds to validate the methodology. Seed regions of reading network: The seed regions for the reading network were selected based on areas known to activate during a reading task, including Left Supramarginal Gyrus (L-SMG), Left Superior Temporal Gyrus (L-STG), Left Fusiform Gyrus (L-FG), Left pars Opercularis (L-pOP). Statistical tests: We conducted F-test to evaluate goodness of fit for 3D modeling, and effect size ‘r’ and ICC to extract highly connected and highly reproducible rsFC maps at group level.Results
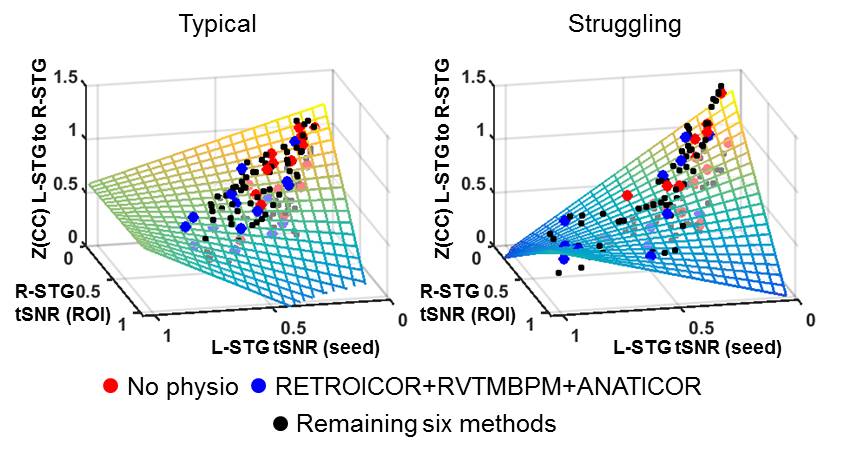
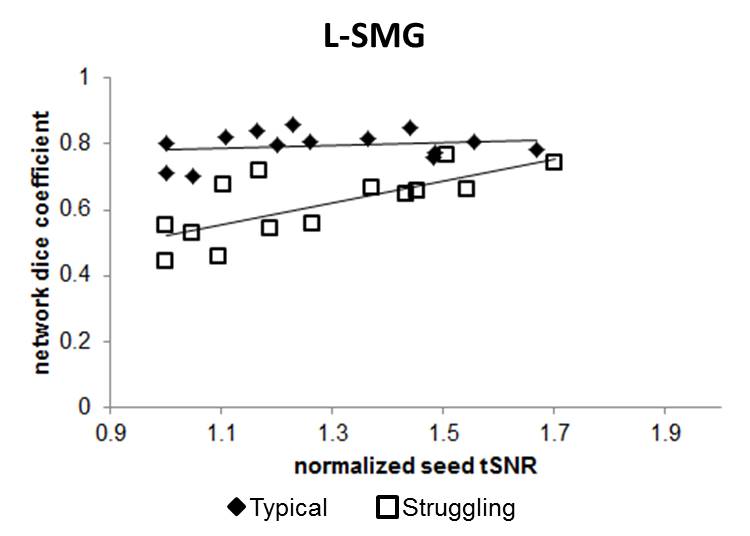
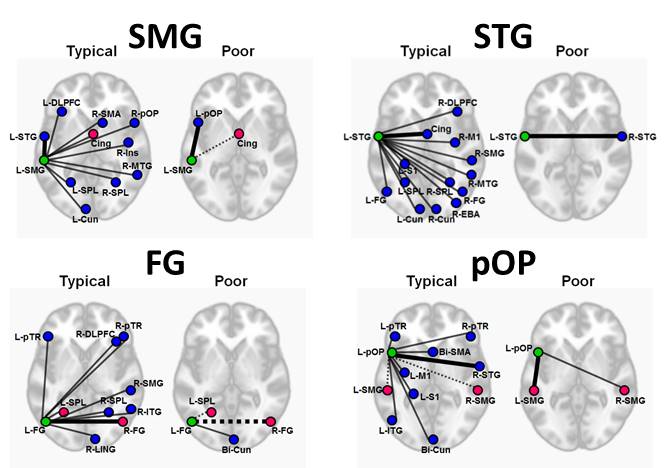
For all LH seed regions, the No Physio pre-processing resulted in the lowest seed tSNR, and the RETROICOR+RVTMBPM+ANATICOR pre-processing resulted in the highest tSNR. Via modeling, we found that the improved tSNR after PN correction from both the seed region and region of interest (ROI) reduced the L-STG to R-STG connectivity in both typical (F=588, p<0.00001) and struggling readers (F=376, p<0.00001) (Figure 1). We also found that higher tSNR results in increased ICC. Furthermore, PN does not affect the rsFC specificity in typical readers, while in struggling readers, the most stringent PN correction helps in increasing the rsFC specificity (Figure 2). Since the highest tSNR, ICC, and DC could be achieved via RETROICOR+RVTMBPM+ANATICOR, we chose to compare the reproducible networks with this method of pre-processing. As seen in Figure 3, the typical readers have a large number of highly connected (effective r > 0.25), and highly reproducible (ICC > 0.65) inter-hemispheric connections, whereas the struggling readers have few or no inter-hemispheric connections, and a reduced number of reproducible connections. Since language and reading is left lateralized, for typical readers, as expected, the RH seeds did not show the same network as the LH seeds.Discussion and Conclusions
PN
artificially inflates the correlation strength leading to erroneous
interpretation. Hence a combination of multiple methodologies to correct for
such noise leads to improved signal fluctuations (tSNR) that culminates in higher
specificity and sensitivity to neuronal fluctuations that are closer to
actual ground truth. Applying our methodologies to a ‘reading’ model, we show
that, irrespective of session, correction for PN results in meaningful
discrimination of reading networks between typical and struggling readers.Acknowledgements
No acknowledgement found.References
[1] Bianciardi M et.al Magn Reson Imaging 2009. [2] Chang C et.al Neuroimage 2009. [3] Birn RM et.al Brain Connect 2014. [4] Birn RM et.al Neuroimage 2006. [5] Murphy K et.al Neuroimage 2009. [6] Glover GH et.al Magn Reson Med 2000. [7] Jo HJ et.al Neuroimage 2010. [8] Pinter D et.al PLOS ONE 2016. [9] Shrout P et.al Psychol. Bull. 1979.Figures