5364
Repetition effects in early blind adults revealed by fMRI adaptation1Integrated Program in Neuroscience, McGill University, Montreal, QC, Canada, 2Linguistics Department, Université du Québec à Montréal, Montréal, QC, 3School of Communication Sciences and Disorders, McGill University, Montréal, QC, Canada, 4Haskins Laboratories, New Haven, CT, United States
Synopsis
Behavioural and fMRI studies have revealed enhanced auditory abilities and differences in the brain organization of early blind individuals compared to sighted. We used fMRI to assess cortical interactions associated with speech processing of repeated and novel stimuli. Although the in-scanner behavior for the blind and sighted groups were comparable, there were a number of cortical activation and deactivation differences. Additionally, the blind participants were better at a post-scan recall task. Cross-modal recruitment of occipital areas was found in the blind participants. They also showed widespread repetition-enhancement effects, suggesting that additional attention mechanisms contribute to their enhanced auditory word encoding abilities.
Introduction
In the past two decades, a great deal of scholarly
literature has established that the functional organization of the brain of early
blind people (EB) differs from that of sighted individuals (SI). As for
perceptual processing, fMRI studies reveal that long-term cross-modal plasticity
is characterized by the recruitment of different cerebral networks. For speech
processing, in addition to auditory areas, visual and audiovisual areas are
more activated in EB 1-5. Differences
in perceptual processing between EB and SI are also observable at the
behavioral level 3,6-10. The EB
constitute a particularly interesting model to understand the developmental plasticity
and reorganization characteristics of processing networks and their behavioral manifestations.
To get a more comprehensive view of the underlying mechanisms associated with
enhanced auditory processing we explored another aspect of cerebral function; neural
adaptation (NA). NA is a phenomenon in which the repetition of a stimulus triggers
a change in the response to the stimulus, upon repeated presentation. This
mechanism is well known in neurophysiology, where these adaptation mechanisms
occur at the level of the single neuron or group of neurons. FMRI adaptation
protocols have highlighted similar mechanisms at the scale of BOLD signal
measurement. Repetition-suppression (RS) is associated with a reduction in the
BOLD response in areas processing a repeated stimulus, and the opposite
response, repetition-enhancement (RE) is associated with stimulus recognition,
learning, attention, expectation and explicit memory 11.
In a previous study 2 RS was observed in auditory areas of both EB and SI during
passive perception of repeated vowel sounds. In EB, we observed additional
areas of RS that included visual (occipital) and audiovisual (superior parietal
lobule) processing regions. In the current study, words and pseudowords were
presented repeatedly. In contrast to our previous study, the repetitions were
presented discontinuously from one another and participants were engaged in an
active lexical decision task.
Methods
Fourteen EB participants and fourteen SI participated in the study. Functional volumes were acquired using a sparse sampling design to avoid acoustic scanner noise12. Stimuli were presented either once or 6 times throughout the experiment. After each trial, participants made lexical judgments (whether the stimulus item existed or not). After the MRI session, participants were tested on the material they heard during the MRI session.Results and Conclusion
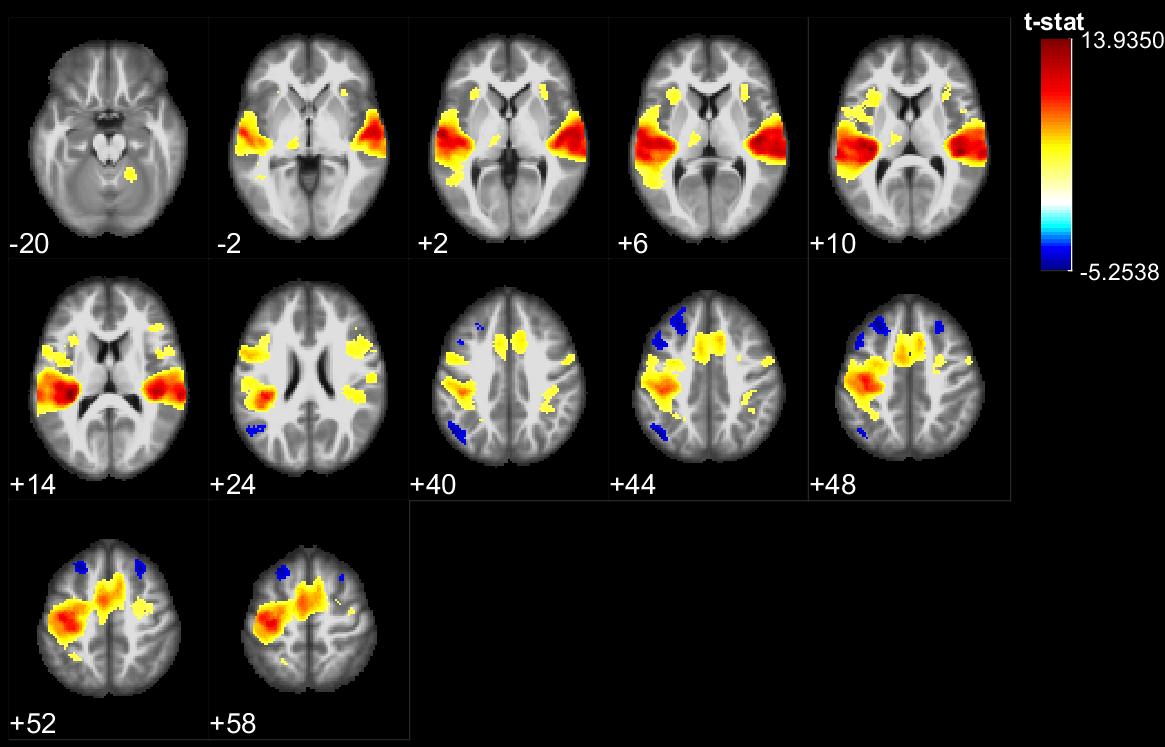
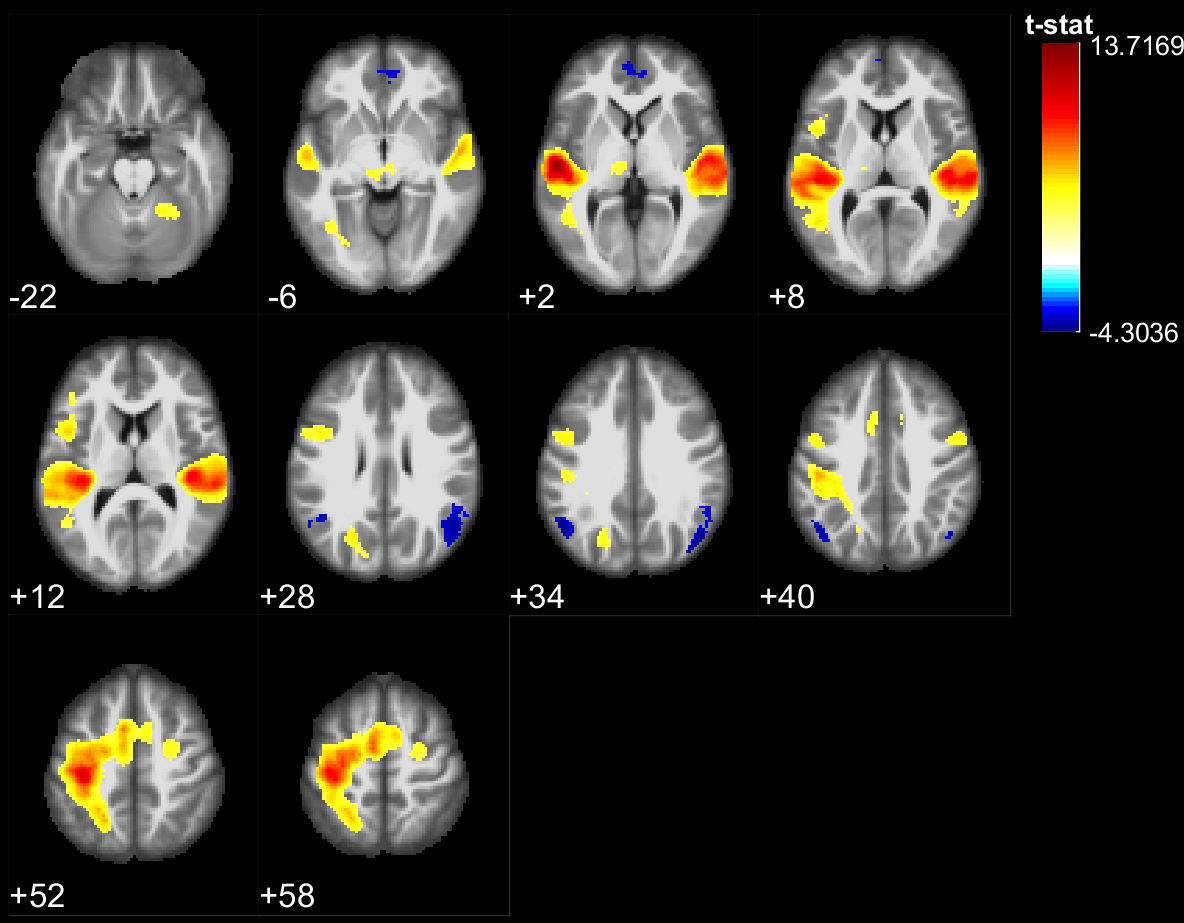
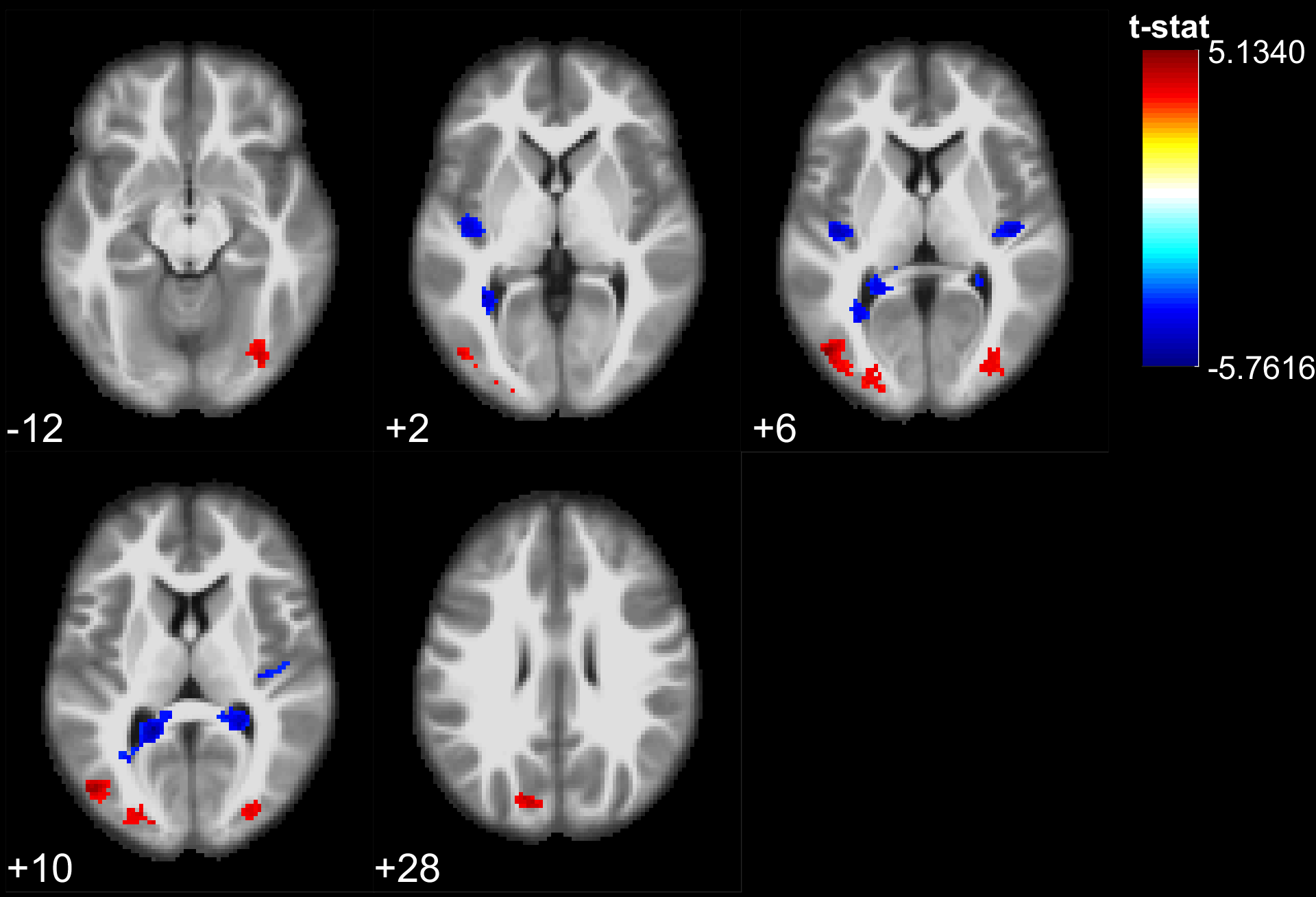
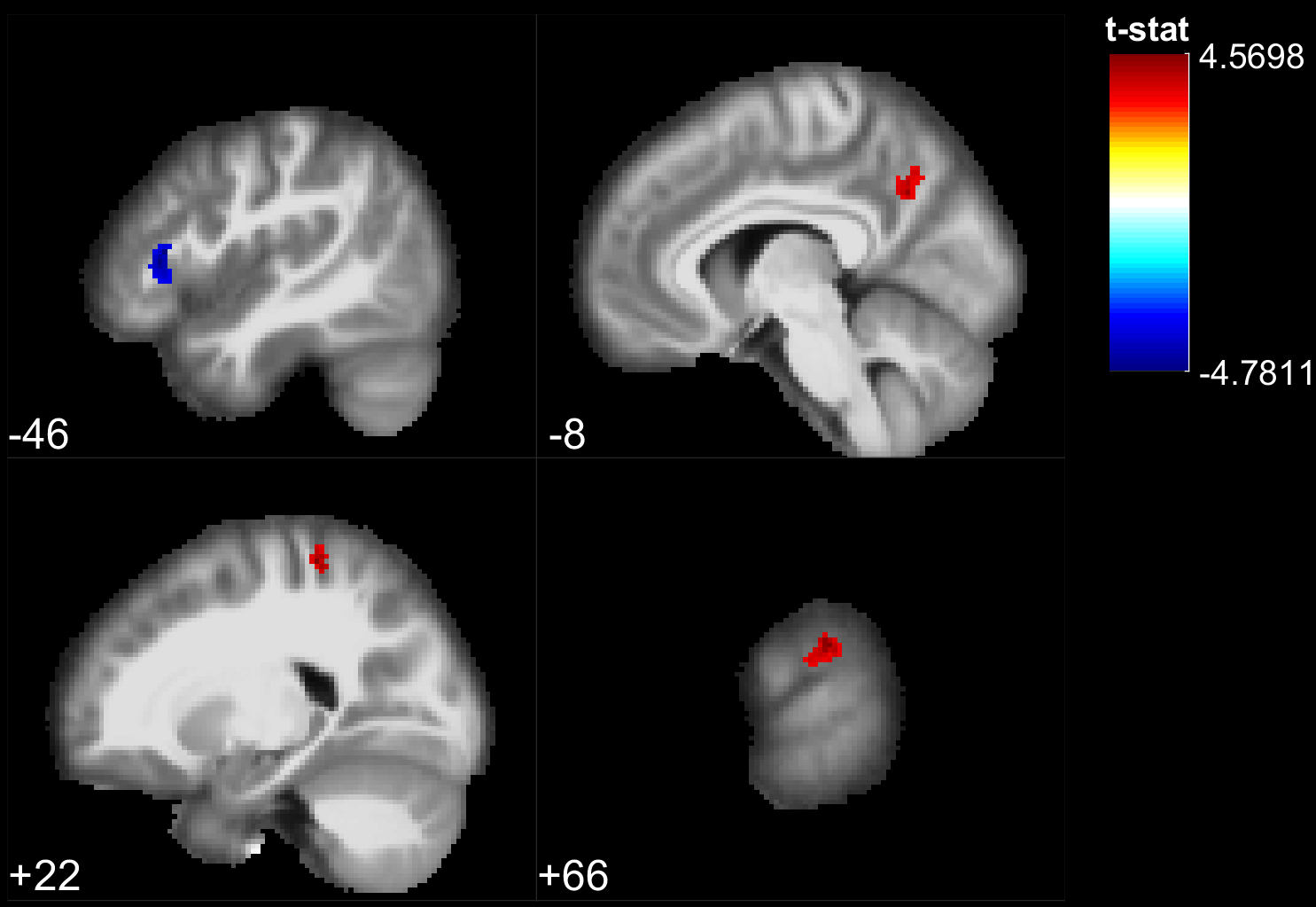
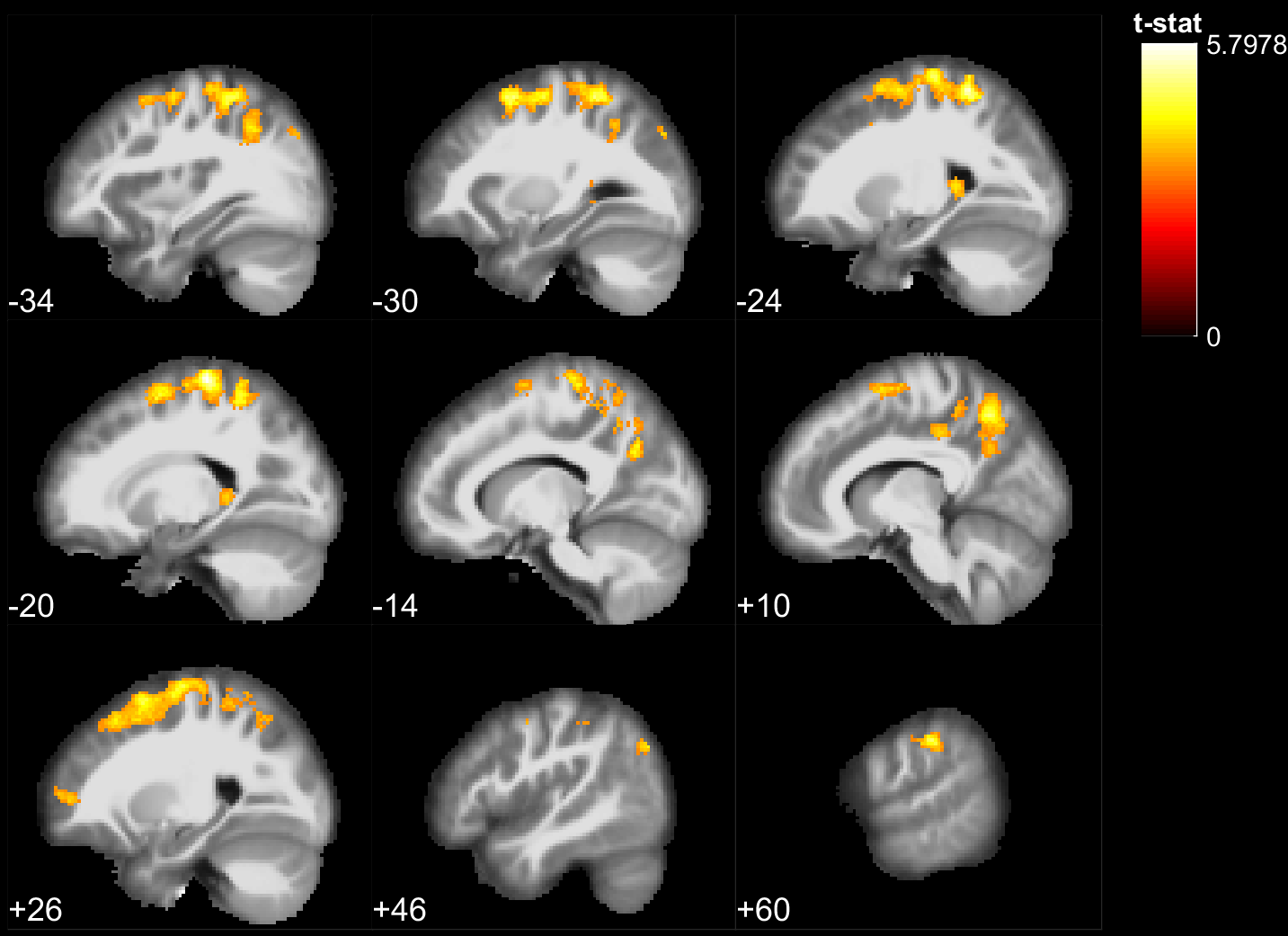
The results of the lexical decision task show that words were answered faster than pseudowords (F(1,28) = 66.5, p<10^-5), that there was a repetition priming effect (F(1,28) = 207.2, p<10^-5) and that there was no significant difference between the groups reaction time (F(1,28) = 0.06, p>0.8). However, the EB were significantly better to recall if a stimulus had been presented or not during the experiment (F(1, 28) = 8.22, p=.0078). During stimulus perception, temporal, parietal and frontal areas traditionally involved in lexical decision tasks were activated in both groups (Fig 1,2). In addition the EB showed cross-modal activity in the occipital lobe (Fig 1). Two-sample t-tests showed no significant difference between the groups for words. However, for pseudowords the EB activated significantly more than SI, portions of the precuneus, cuneus, fusiform gyrus, inferior occipital gyri, middle occipital gyri and superior occipital gyri. The SI had significantly more activity than EB in bilateral clusters around the hippocampus, as well as in the insula and Heschl’s gyrus for unique pseudowords (Fig 3). Regarding adaptation effects, the controls showed repetition-suppression effects in the left inferior frontal gyrus, and repetition-enhancement in the right SMG, left precuneus and right postcentral gyrus (Fig 4). The blind did not show any repetition-suppression effect, however they showed widespread repetition-enhancement effects in portions of the parietal lobe and frontal lobe including the angular gyrus, the precuneus, the pre and post-central gyri, as well as in the middle occipital gyrus, and left hippocampus (Fig 5). Our first study indicated that repetition-suppression was occurring during the passive perception of repeated sounds, and that these mechanisms were also present cross-modally in EB. Here, we show widespread repetition-enhancement effects during the active processing of words and pseudowords in EB only. The presence of repetition-enhancement effects seems to indicate that during an active task, additional phenomenon, such as stimulus recognition, expectation and more generally attention mechanisms could contribute to the enhanced ability of blind participants to process auditory stimuli. Taken together the results indicate that the long-term neuroplastic reorganization occurring in the blind brain not only involves cross-modal plasticity of sensory areas but also a functional reorganization of auditory attention mechanisms that contribute to the enhanced auditory encoding of sounds.Acknowledgements
Research supported by a Discovery Grant from NSERC (awarded to V. Gracco).
References
1 Roder, B., Stock, O., Bien, S., Neville, H. & Rosler, F. Speech processing activates visual cortex in congenitally blind humans. Eur J Neurosci 16, 930-936 (2002).
2 Arnaud, L., Sato, M., Menard, L. & Gracco, V. L. Repetition suppression for speech processing in the associative occipital and parietal cortex of congenitally blind adults. PloS one 8, e64553, doi:10.1371/journal.pone.0064553 (2013).
3 Dietrich, S., Hertrich, I. & Ackermann, H. Ultra-fast speech comprehension in blind subjects engages primary visual cortex, fusiform gyrus, and pulvinar -- a functional magnetic resonance imaging (fMRI) study. BMC Neurosci 14, 74, doi:10.1186/1471-2202-14-74 (2013).
4 Bedny, M., Pascual-Leone, A., Dravida, S. & Saxe, R. A sensitive period for language in the visual cortex: distinct patterns of plasticity in congenitally versus late blind adults. Brain Lang 122, 162-170, doi:10.1016/j.bandl.2011.10.005 (2012).
5 Bedny, M., Pascual-Leone, A., Dodell-Feder, D., Fedorenko, E. & Saxe, R. Language processing in the occipital cortex of congenitally blind adults. Proc Natl Acad Sci U S A 108, 4429-4434, doi:10.1073/pnas.1014818108 (2011).
6 Hugdahl, K. et al. Blind individuals show enhanced perceptual and attentional sensitivity for identification of speech sounds. Brain Res Cogn Brain Res 19, 28-32 (2004).
7 Menard, L., Dupont, S., Baum, S. R. & Aubin, J. Production and perception of French vowels by congenitally blind adults and sighted adults. J Acoust Soc Am 126, 1406-1414, doi:10.1121/1.3158930 (2009).
8 Hertrich, I., Dietrich, S., Moos, A., Trouvain, J. & Ackermann, H. Enhanced speech perception capabilities in a blind listener are associated with activation of fusiform gyrus and primary visual cortex. Neurocase 15, 163-170, doi:10.1080/13554790802709054 (2009).
9 Hertrich, I., Dietrich, S. & Ackermann, H. Tracking the speech signal--time-locked MEG signals during perception of ultra-fast and moderately fast speech in blind and in sighted listeners. Brain Lang 124, 9-21 (2013).
10 Hertrich, I., Dietrich, S. & Ackermann, H. How can audiovisual pathways enhance the temporal resolution of time-compressed speech in blind subjects? Frontiers in psychology 4, 530, doi:10.3389/fpsyg.2013.00530 (2013).
11 Segaert, K., Weber, K., de Lange, F. P., Petersson, K. M. & Hagoort, P. The suppression of repetition enhancement: a review of fMRI studies. Neuropsychologia 51, 59-66 (2013).
12 Perrachione, T. K. & Ghosh, S. S. Optimized design and analysis of sparse-sampling FMRI experiments. Front Neurosci 7, 55, doi:10.3389/fnins.2013.00055 (2013).
Figures