5363
A BOLD analysis of the olfactory perception system in the mouse whole brain, using independent component analysisHirotsugu Funatsu1, Fuyu Hayashi1, Sosuke Yoshinaga1, Mitsuhiro Takeda1, Naoya Yuzuriha1, Shunsuke Kusanagi1, and Hiroaki Terasawa1
1Department of Structural BioImaging, Faculty of Life Sciences, Kumamoto University, Kumamoto, Japan
Synopsis
A BOLD analysis can trace real-time odor responses at high spatial resolution. Rodents have well-developed olfactory systems, and thus BOLD studies have mainly been performed in rodents. However, mice have smaller brains and are more susceptible to peripheral hemodynamic changes than rats, which make it harder to obtain sufficient BOLD signals. This study revealed the real-time odor response in the mouse whole brain by the BOLD experiment with periodical odor stimulations using a high-sensitivity cryogenic probe and the high-detectability analytical method, Independent Component Analysis (ICA). The signal changes of the activation sites corresponded to the human canonical hemodynamic response function.
Introduction
Olfaction is one of the most important perceptions for animals to receive information from their environments. When odorant compounds are inhaled, the olfactory bulb (OB) shows unique activation patterns depending on the types of odors. However, our understanding of the odor-stimulated activation patterns over the whole brain is still limited, and thus the means by which the brain discriminates between different odors remain to be elucidated [1]. A BOLD analysis is the main method that monitors real-time odor responses at high spatial resolution in studies of rodents, which have well-developed olfactory systems. Among rodents, the number of odor fMRI studies of mice is smaller than that of rats. Some studies have been reported for the mouse OB [2], but the number of fMRI studies for the mouse whole brain is much lower than that for the mouse OB [3]. One reason for this is that the mouse brain is smaller and thus the BOLD analysis is more likely to suffer from peripheral hemodynamic changes, which make it harder to obtain sufficient fMRI signals [4]. Thus, a robust procedure is required to detect odor responses in the mouse whole brain. Independent component analysis (ICA) is a high-detectability analytical method that can identify periodically activated brain regions. It can detect stimulation-evoked responses by applying periodical stimulations and tracing the MR signal transition components that have the same frequency as the stimulation frequency [5]. ICA was successfully employed in some fMRI studies of rats [6, 7]. Here, we applied ICA to the BOLD experiment in which odor stimulation was periodically applied to mice during GRE-EPI scans at high-temporal resolution. In addition, a cryogenic probe was used to improve the detectability of the BOLD response. The aim of this study is to establish a method that can detect odor responses in the mouse whole brain and to reveal the mechanism of odor discrimination, by using the above-mentioned elemental techniques.Methods
MRI experiments were performed with a 7.0 Tesla Bruker BioSpec 70/20 scanner and a mouse brain 2-channel phased array surface cryogenic coil (Bruker BioSpin). Mice (male C57BL/6, 8–10 weeks old) were anesthetized with urethane (i.p. 1.5 g/kg initial; 0.1 g/kg/hr supplemental) and GRE-EPI images were acquired: TR/TE=4000/21.3 ms; FOV=1.28×1.28 cm2; matrix=64×64; resolution=200×200 µm2; slice thickness=400 µm; slices=30; NEX=1; flip angle=90°. In the middle of the scans, isoamyl acetate (IAA) vapor was mixed with the air flow and delivered to the mouse nose. The odor stimulation consisted of 24 tasks of 5 sec ON and 55 sec OFF. The images were analyzed by ICA with the FSL software. The MR signal transition components with the same frequency as the odor stimulations were selected to identify the activated regions in the mouse brains.Result
As a result of the ICA of the MR data, the components that had the 16.6 mHz signal frequency were detected in the piriform cortex (Fig.1A), the dorsal OB (Fig.1B) and a wide range region that includes the posterior lateral hypothalamus (PLH), the medial part of the extended amygdala (EAM) and the central part of the extended amygdala (EAC). In each region, an increase of the signal intensity was observed about 8 sec after the stimulation on the signal time course, which was generated by summing the intensity profiles of the 24 tasks (Fig.2). The observed delay was consistent with the human canonical hemodynamic response function and the results of a previous study, in which the rat OB was repeatedly stimulated by IAA [8].Discussion
The piriform cortex, which belongs to an olfactory pathway and thus was expected to be activated by odor stimulation, was actually detected in this study (Fig.1A). Consistent with our results in mice, the activation in the dorsal OB was reported in IAA-stimulated rats [2, 8], suggesting the similar activation pattern by IAA (Fig.1B). The PLH was also activated (Fig.1C), which may indicate the feeding center activation [9] by the banana-like odor of IAA. When the 24 tasks were divided into the first half and the second half, the BOLD response summed over the second half was smaller than that summed over the first half, possibly due to the effect of desensitization [2].Conclusion
This study revealed the
real-time activation of the mouse whole brain by odor stimulation, based on the
signal intensity analysis with ICA. We
plan to examine other anesthetic agents, to explore brain
activation in more detail. We also plan to analyze and compare the activation
of brain regions evoked by other odorants, to reveal the mechanism of odor
discrimination.Acknowledgements
This work was supported in part by Grants-in-Aid for Scientific Research (B) (JP15H04489) (to H.T.) and Scientific Research (C) (JP26450129) (to S.Y.) from the Japan Society for the Promotion of Science.References
[1] Touhara, K. and Vosshall, L.B., Annu. Rev. Physiol. 71, 307–332 (2009), [2] Xu, F. et al., Proc. Natl. Acad. Sci. USA 100, 11029–11034 (2003), [3] Pain, F. et al., Cell. Mol. Life Sci. 68, 2689–2709 (2011), [4] Ielacqua, G.D. et al., Proc. Intl. Soc. Mag. Reson. Med. 23, 2037 (2015), [5] Sinieral, J.D. et al., Proc. Intl. Soc. Mag. Reson. Med. 24, 1716 (1999), [6] Berge, N. et al., PLOS ONE. 10, 7 (2015), [7] Yuzuriha, N. et al., Proc. Intl. Soc. Mag. Reson. Med. 24, 1722 (2016), [8] Martin, C. et al., NeuroImage 36, 1288–1293 (2007), [9] Li, J. et al., Br. J. Pharmacol. 171, 332–350 (2014)Figures
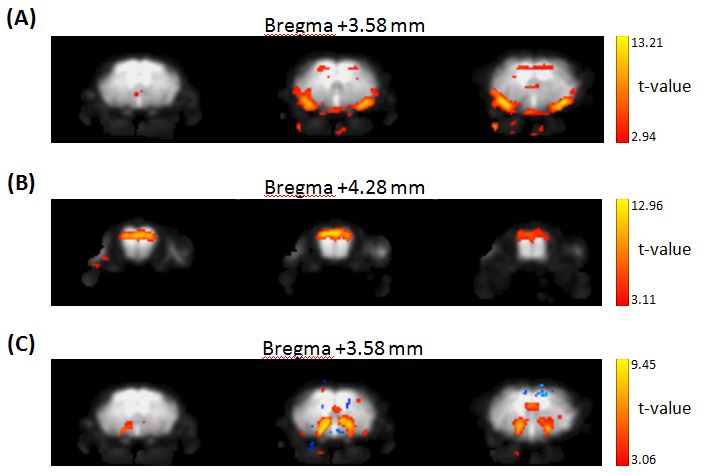
Fig. 1 MR signal transition
components that have the same frequency as the 24 tasks of odor stimulations in
the piriform cortex (A), the dorsal OB (B), the wide range region of the posterior lateral
hypothalamus (PLH), the medial part of extended amygdala (EAM) and the central part of
extended amygdala (EAC) (C).
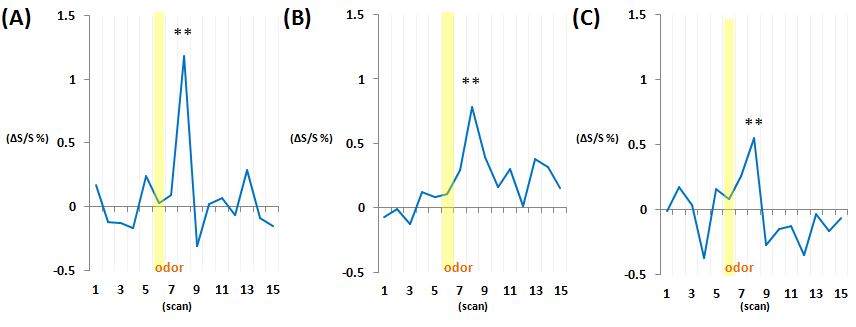
Fig. 2 Signal time courses that were generated by
summing the 24 odor tasks. Yellow
squares show the duration of the odor stimulations in the piriform cortex (A), the dorsal
OB (B), the wide range region of PLH, EAM and EAC (C). (**, p < 0.01)