4070
Environmental enrichment changes neuroanatomy more than exercise and does not require CREB1Mouse Imaging Centre, Hospital for Sick Children, Toronto, ON, Canada, 2Dept. of Medical Biophysics, University of Toronto, Toronto, ON, Canada, 3Hospital for Sick Children, Toronto, ON, Canada, 4Dept. of Psychology, University of Toronto, Toronto, ON, Canada
Synopsis
Human and rodent imaging studies show that experience can lead to task-specific increases in brain structure, but the cellular and molecular basis of these changes is unknown. Here, we test the hypothesis that the these volume changes depend on the same signaling pathways required for learning and memory formation. We exposed mice lacking CREB (a transcription factor critical for memory) to an enriched housing environment, and imaged their brains longitudinally over two weeks. Surprisingly, enrichment altered the neuroanatomy of all mice, regardless of CREB genotype. These findings provide evidence that the volume changes are due to CREB-independent processes.
Introduction
Human imaging studies show that experience can lead to anatomical brain changes at a scale detectable with MRI. For instance, learning to juggle or to navigate a complex environment can alter the volume of task-specific brain regions1-3. However, the cellular mechanisms and signalling pathways underlying these volume changes are poorly understood.
Mouse imaging studies are ideal for probing the basis of these volume changes: mice are genetically similar to humans, have easily editable genomes, and display low genetic variability4. In mice, environmental enrichment is a well-established paradigm to investigate the effect of experience on the brain. An enriched environment provides more stimulation than standard housing, and can include running wheels, tunnels, and increased social interaction. Enrichment has been associated with a variety of cellular alterations to the brain, including increases in dendritic branching, neurogenesis, and changes in cortical thickness5,6. Additionally, enrichment is associated with increases in hippocampal volume that can be detected with mouse MRI7. However, the link, if any, between cellular and volume changes is unknown.
One hypothesis is that the neuroanatomical changes induced by enrichment are due to learning to navigate a spatially complex environment. Given this, we hypothesized that we could block the volume changes by targeting the signalling pathways implicated in learning and long term memory formation (LTM). The transcription factor CREB is critical for LTM and regulates many cellular processes stimulated by enrichment, including dendritic growth8,9. Here, we used CREB mutant mice to ask whether the neuroanatomical changes induced by environmental enrichment require CREB-dependent signaling (Figure 1). An alternative hypothesis is that the enrichment effects are due to exercise, not spatial learning. We tested this by comparing volume changes induced by enrichment to exercise alone in wildtype and CREB mutant mice.
Methods
Enrichment and Imaging: Adult (~8 weeks) male and female CREBɑδ mutant mice (CREBɑδ+/+, CREBɑδ+/-, CREBɑδ-/- on a B6/129 background) were imaged at baseline on a 7T scanner using in vivo manganese-enhanced MRI (90 μm resolution, TR=26ms, TE=5.37ms, 1 hour 52 min. scan time). Mice were then placed into group Enriched Housing, group Standard Housing, individual exercise-only housing, or individual standard housing (N=12-28 per genotype and housing condition) and imaged after 48 hours, 8 days, and 16 days. The enriched environment featured 3 levels of tubes and tunnels, nesting material, and a running wheel7(Figure 2). The exercise-only cages contained only nesting material and a running wheel.
Image analysis: The images for each mouse at each timepoint were aligned to create subject-specific averages, which were then aligned to create a study average7,10. This study average was then registered to an atlas and used with each image’s deformation fields to compute the volumes of 62 distinct brain structures for each image11,12. The Jacobian determinant of the deformation field was computed as a measure of local expansion or contraction at each voxel. The effect of genotype and housing treatment on structure and voxel volume was assessed using linear regression. Multiple comparisons were controlled used the False Discovery Rate13.
Results
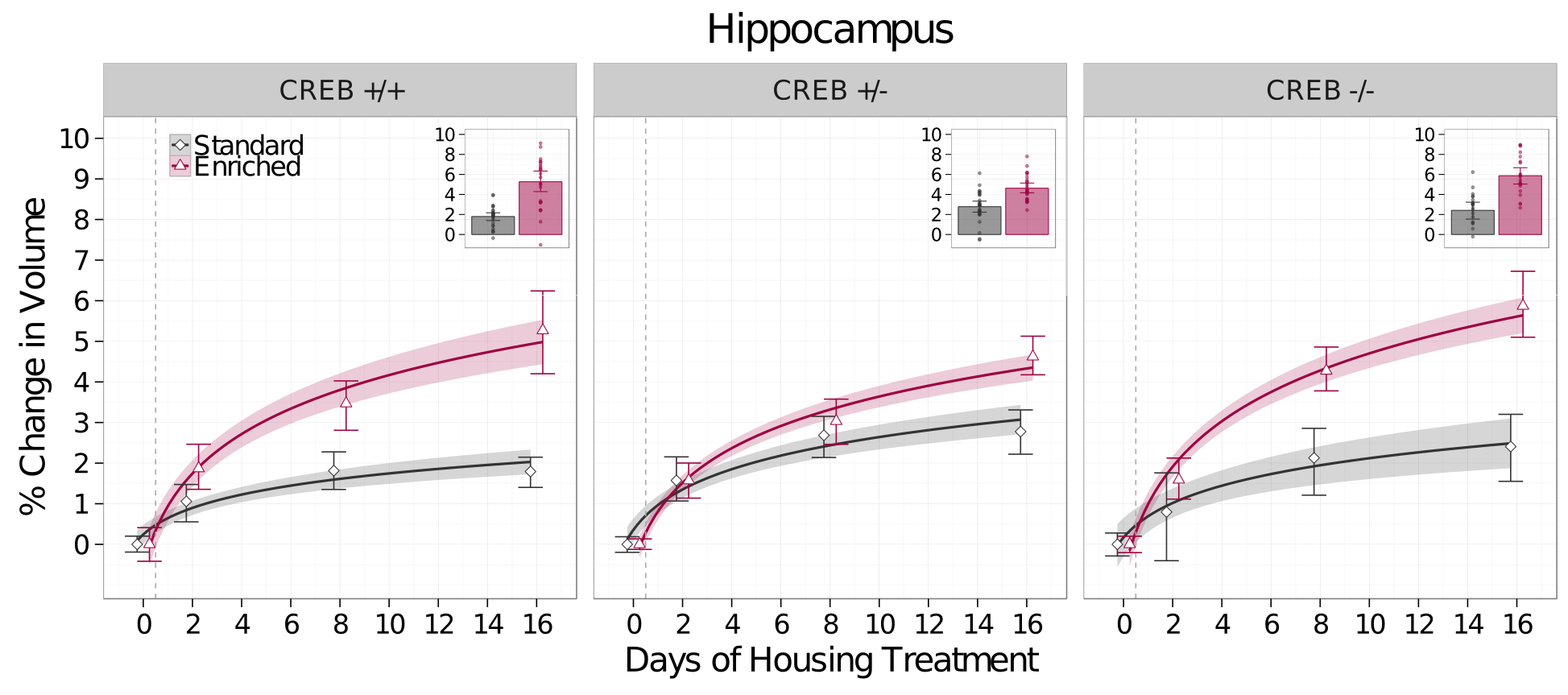
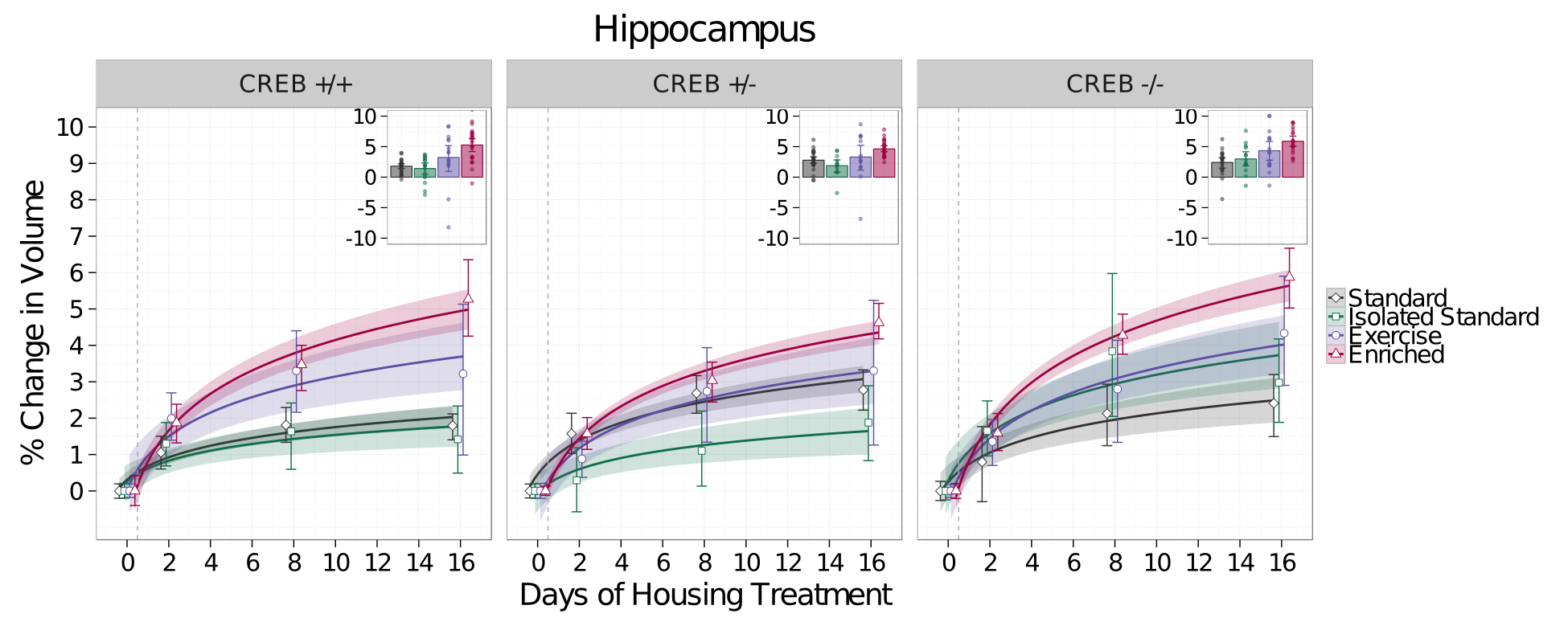
There was a significant increase in the growth rate of the hippocampus in Enriched mice compared to Standard housed mice regardless of CREB genotype (t=6.08,p<0.0001,Figure 3). These results suggested an alternative model - that volume changes are mediated by the additional exercise the mice experience in the enriched housing, not spatial learning, and that they therefore may be CREB-independent. To test this, an additional cohort of mice was placed in individual cages with running wheels only, and imaged at the same timepoint as described above. To control for the effects of social isolation, a separate cohort of mice was also placed in individual standard housing. Exercise also increased the rate of growth of the hippocampus compared to Standard housed mice independent of CREB genotype (t=2.31,p=0.01); however the rate of growth in Enriched mice was higher still than that of Exercise-only mice (t=2.822,p=0.005, Figure 4).Discussion and Conclusions
Surprisingly, environmental enrichment increases the volume of the hippocampus regardless of CREB genotype. Moreover, the effects of enrichment on hippocampal volume were only partially mediated by exercise. These results led us to two conclusions: First, while the exercise-component of enrichment does alter brain anatomy, the other factors (spatial learning, and the interaction effects between learning, exercise, and social interaction), result in additional increases in hippocampal volume. Secondly, contrary to our hypothesis, the neuroanatomical changes seen with MRI are mediated by CREB-independent processes or only residual CREB function is required. Future experiments will investigate whether the cellular impact of enrichment is the same across all CREB genotypes, and investigate CREB-independent mechanisms that may mediate the volume changes.Acknowledgements
The authors wish to thank Matthijs van Eede and Ben Darwin for computing support. Funding for this work was provided by the Canadian Foundation for Innovation (CFI), the Canadian Institute for Health Resesarch (CIHR), and an Ontario Graduate Scholarship.References
1. Zatorre, R. J., Fields, R. D. & Johansen-Berg, H. Plasticity in gray and white: neuroimaging changes in brain structure during learning. Nat Neurosci 15, 528–536 (2012).
2. Draganski, B. et al. Neuroplasticity: Changes in grey matter induced by training. Nature 427, 311–312 (2004).
3. Maguire, E. A. et al. Navigation-related structural change in the hippocampi of taxi drivers. Proc Natl Acad Sci U S A 97, 4398–4403 (2000). 4. Henkelman, R. M. Systems biology through mouse imaging centers: experience and new directions. Annual Review of Biomedical Engineering 12, 143–166 (2010).
5. van Praag, H., Kempermann, G. & Gage, F. H. Neural consequences of environmental enrichment. Nat Rev Neurosci 1, 191–198 (2000).
6. Diamond, M. C., Ingham, C. A., Johnson, R. E., Bennett, E. L. & Rosenzweig, M. R. Effects of environment on morphology of rat cerebral cortex and hippocampus. J. Neurobiol.7, 75–85 (1976).
7. Scholz, J., Allemang-Grand, R., Dazai, J. & Lerch, J. P. Environmental enrichment is associated with rapid volumetric brain changes in adult mice. NeuroImage 109, 190–198 (2015).
8. Barco, A., Jancic, D. & Kandel, E. R. CREB-dependent transcription and synaptic plasticity. Transcriptional Regulation by Neuronal … (2008).
9. Bourtchuladze, R. et al. Deficient long-term memory in mice with a targeted mutation of the cAMP-responsive element-binding protein. Cell 79, 59–68 (1994).
10. Scholz, J., Klein, M. C., Behrens, T. E. J. & Johansen-Berg, H. Training induces changes in white-matter architecture. Nat Neurosci 12, 1370–1371 (2009).
11. Lerch, J. P., Sled, J. G. & Henkelman, R. M. MRI phenotyping of genetically altered mice. Magnetic Resonance Neuroimaging (2011).
12. Dorr, A. E., Lerch, J. P., Spring, S., Kabani, N. & Henkelman, R. M. High resolution three-dimensional brain atlas using an average magnetic resonance image of 40 adult C57Bl/6J mice. NeuroImage 42, 60–69 (2008).
13. Genovese, C. R., Lazar, N. A. & Nichols, T. Thresholding of statistical maps in functional neuroimaging using the false discovery rate. NeuroImage 15, 870–878 (2002).
Figures