3438
Adipose tissue and ectopic fat responses to an extended fast in healthy male adults1Wolfson Brain Imaging Centre, University of Cambridge, Cambridge, United Kingdom, 2NIHR/WT Clinical Research Facility, Cambridge University Hospitals NHS Foundation Trust, Cambridge, United Kingdom, 3Department of Paediatrics, University of Cambridge, Cambridge, United Kingdom, 4MRC Human Nutrition Research, Cambridge, United Kingdom, 5Department of Musculoskeletal Biology, University of Liverpool, Liverpool, United Kingdom, 6Oxford Centre for Diabetes, Endocrinology and Metabolism, University of Oxford, Oxford, United Kingdom
Synopsis
We have investigated the response of different fat deposits to a 28 h fast in healthy nonobese males and shown significant increases in CH2 IMCL in both the soleus and tibialis anterior muscles, and significant decreases in subcutaneous fat. Increases in TA and SOL IMCL correlated with oleic and palmitic acid FFA concentrations respectively. The compositional change of IMCL during the fast related inversely to the baseline composition, suggesting a selective efflux of unsaturated shorter chain IMCLs for preferential oxidation, which is in agreement with rat biopsy data, and supportive of the idea of IMCL as a flexible lipid store.
Introduction
Triglyceride stores are an important source of fuel during fasting, and studies 1,2 have shown that highly unsaturated and shorter-chained fatty acids are preferentially mobilised and oxidised under these conditions. Here we use 1H MRI and MRS to study the differing adipose tissue and ectopic fat responses to a 28 h fast, and compare these with circulating free fatty acids (FFA) measured by mass spectrometry.Methods
The methyl and methylene protons of intra- and extra-myocellular lipid (IMCL,EMCL) from the tibialis anterior (TA) and soleus (SOL) muscles were measured at 8 and 28 h of fasting in 24 healthy non-obese males (age 34.9±1.8 y, BMI 23.4±0.5 kg.m-2). These were compared with changes in abdominal subcutaneous (SCATabd) and visceral adipose tissue (VAT) volume and calf subcutaneous (SCATleg) and intramuscular (IMF) fat area determined by MRI, and in intrahepatic lipid (IHL) assessed by 1H MRS, as well as circulating FFA. MR was performed on a Siemens 3T Verio, the voxels/slices being carefully relocalised on the second visit. We use the IMCL CH3:CH2 ratio as a marker of the degree of unsaturation combined with chain length, and validate this in agar phantoms of differing fatty acid compositions.
IMCL, EMCL
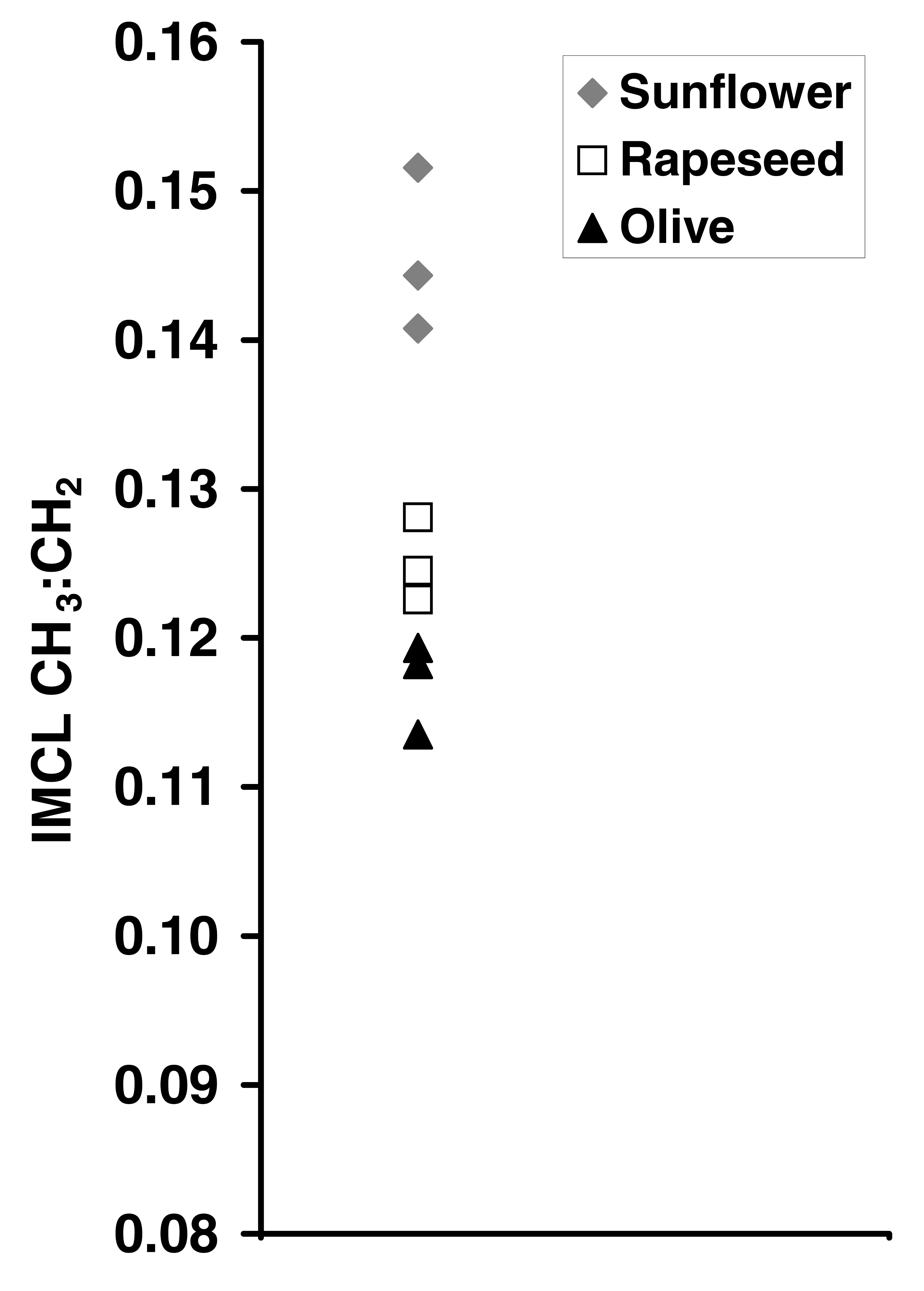
A water-suppressed 1H MR spectrum was obtained using PRESS with TR=5s, TE=35 ms and 64 averages from voxels (cube length 1.3 cm) located within SOL and TA to avoid visible fat on T1-weighted images. Spectra were analyzed in jMRUI 3 and fitted with AMARES 4 algorithm using comprehensive prior knowledge. Agar phantoms simulating IMCL+EMCL 5 were made using olive, rapeseed and sunflower oils. Three 1H MR spectra were acquired from each phantom using the same acquisition and analysis parameters as in vivo.
SCATabd, VAT, SCATleg, IMF, IHL
Abdominal SCATabd and VAT volumes were measured over 6 cm superior-inferior distance from 5 water-suppressed T1-weighted transaxial slices. SCATleg and IMF were assessed from the T1-weighted transaxial slice containing the 1H MRS voxels. IHL was measured using PRESS from a voxel (cube length 1.5 cm) in the posterior aspect of the right lobe of the liver.
FFA
Circulating FFA composition
was assessed using high-resolution mass spectrometry of plasma samples at 12,
16 and 24 h fast, combined as the AUC. FFA concentrations were generated by
proxy and are expressed in arbitrary units.
Results
Non-normal variables were log-transformed prior to statistical tests. Two remaining outliers were eliminated from IHL. Muscle spectra were eliminated if there was no clear distinction between EMCL and IMCL CH2 resonances, yielding 22 and 19 complete pre-post sets for TA and SOL respectively.
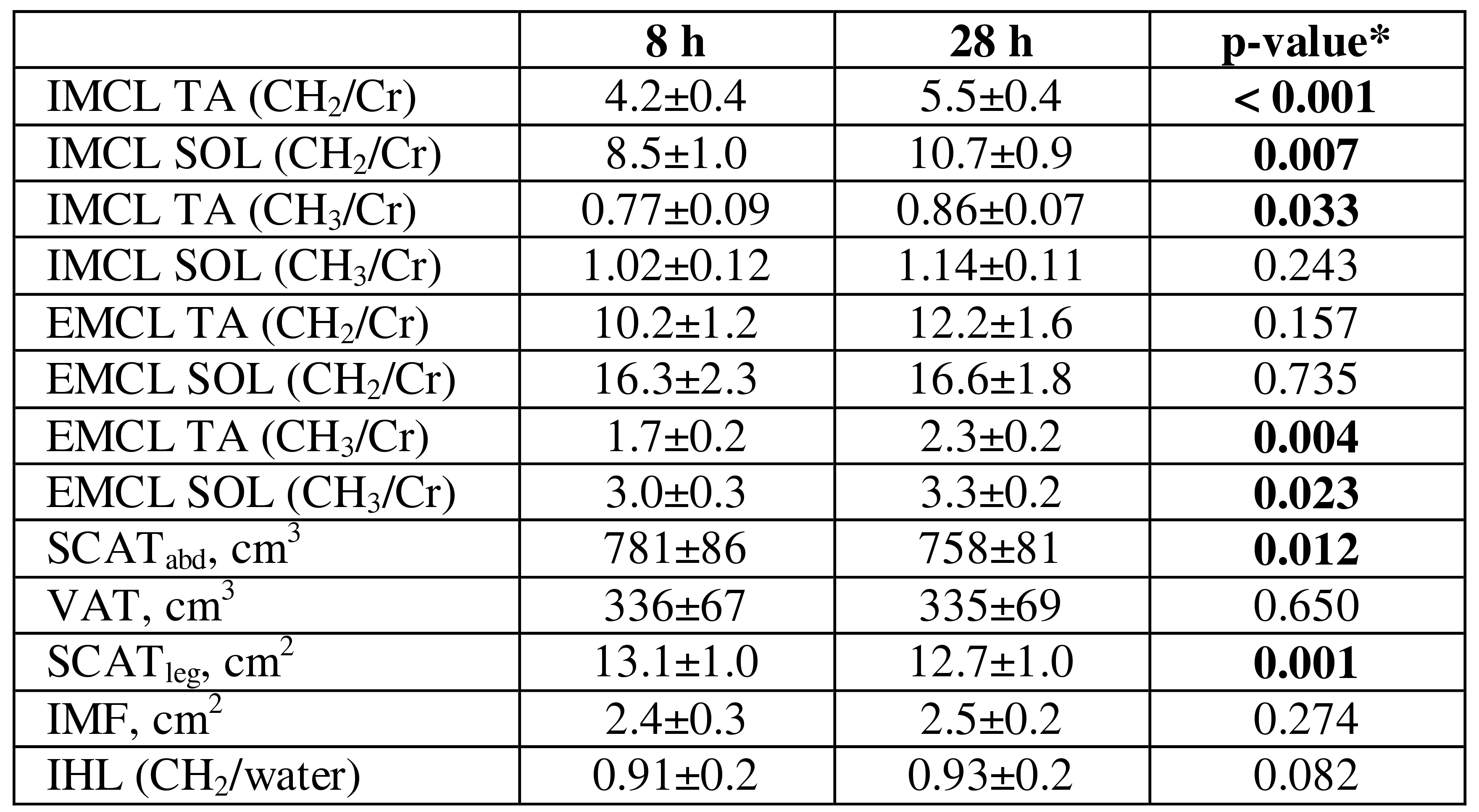
MR measures at 8 and 28 h are shown in Table 1. In both TA and SOL, the difference (Δ) in IMCLCH2 significantly correlated with ΔIMCLCH3 (r=0.71, r=0.88 respectively, p<0.001), as did ΔEMCLCH2 with ΔEMCLCH3 (r=0.88, r=0.86 respectively, p<0.001). ΔSCATleg significantly correlated in TA with CH2 ΔIMCL (r=-0.50,p=0.02) and ΔEMCL (r=-0.46,p=0.03), and in SOL with CH3 (and not CH2) ΔIMCL (r=-0.50,p=0.03).
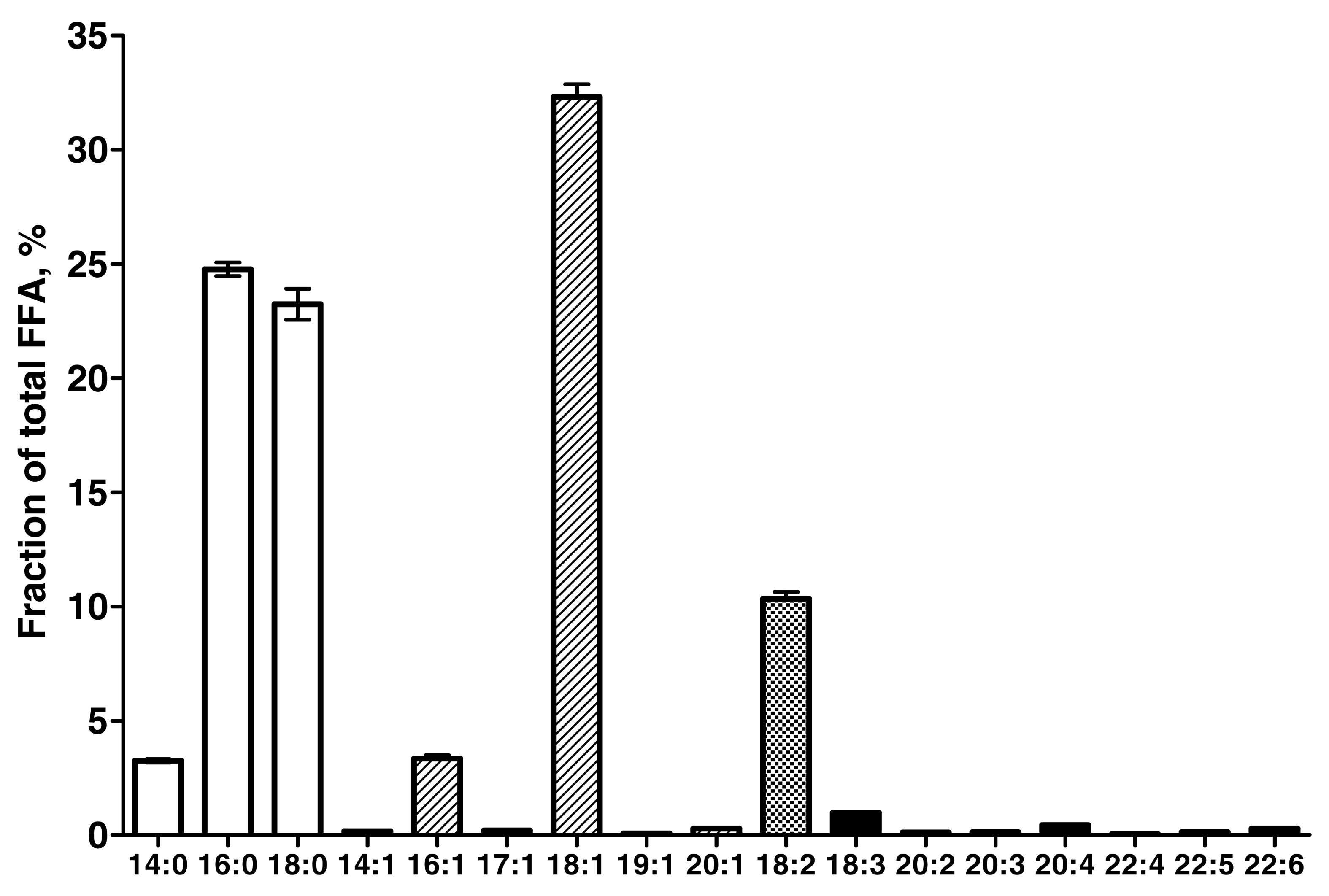
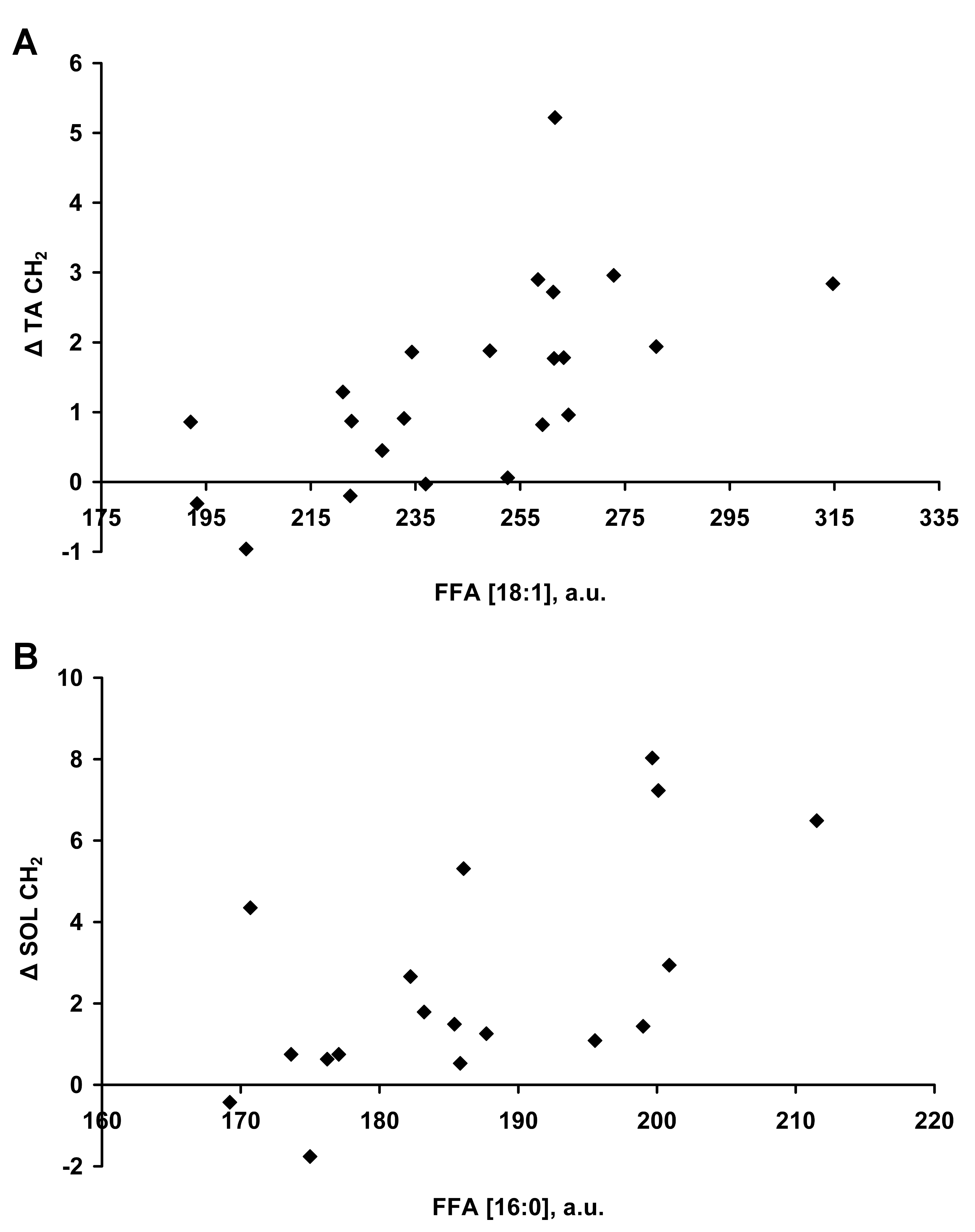
Figure 1 shows the circulating FFA profile during the fast. TA ΔIMCLCH2 significantly correlated with total FFA concentration (r=0.57,p=0.006), in particular total monounsaturated (r=0.60,p=0.003) FFA, specifically [18:1] (r=0.64,p=0.002 Figure 2A), as well as total omega-3 FFA (r=0.45,p=0.04). SOL ΔIMCLCH2 correlated with total FFA (r=0.52,p=0.02), but in contrast to TA this was related specifically to the combined saturated 14:0+16:0, in particular [16:0] FFA (r=0.49,p=0.03, Figure2B).
The
IMCL CH3:CH2 ratio could reliably distinguish the 3
different oil IMCL+EMCL phantoms apart by their relative order (Figure 3). The
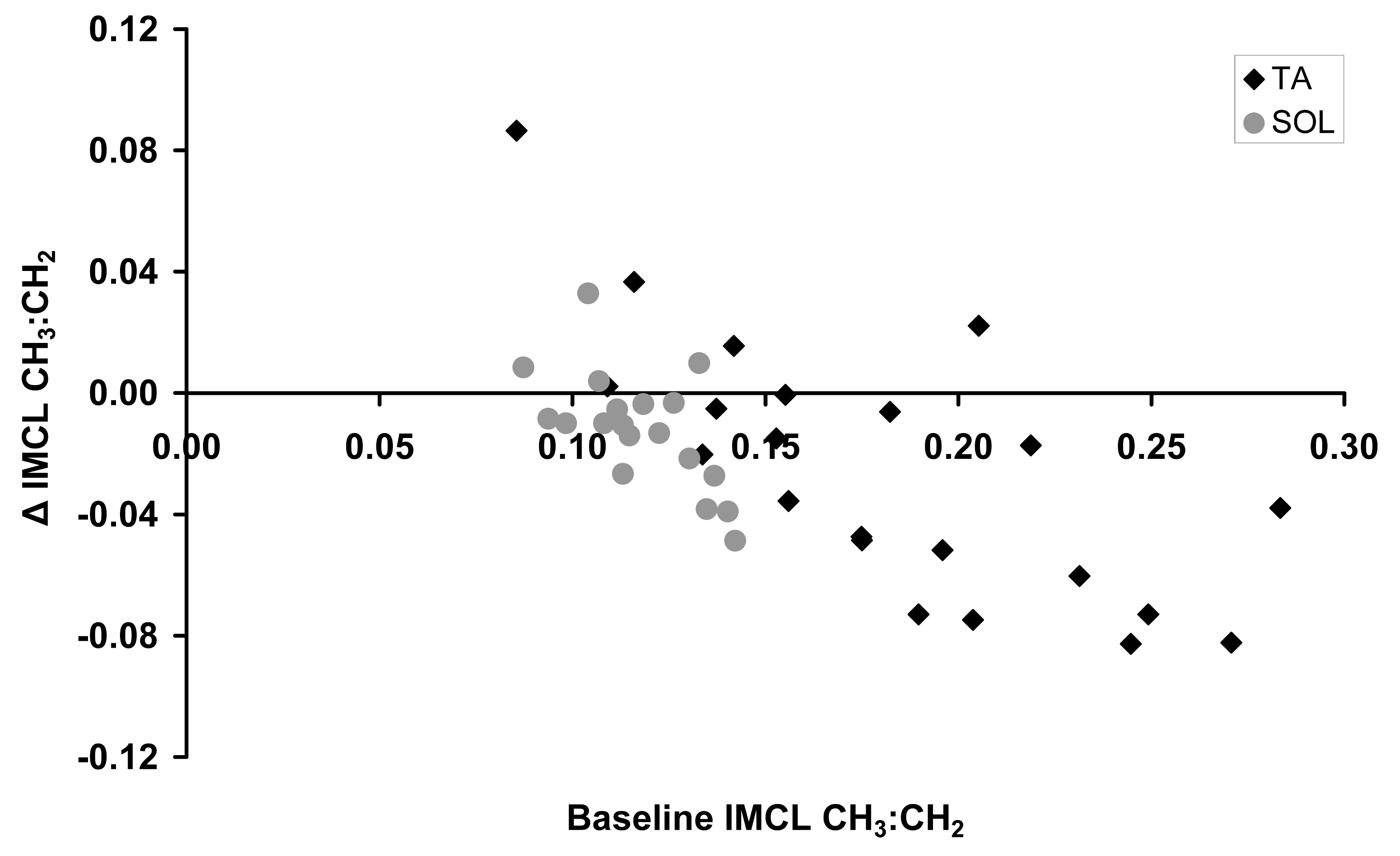
in vivo ΔIMCL CH3:CH2
correlated with the baseline IMCL CH3:CH2 in both TA
(r=-0.73,p<0.001) and SOL (r=-0.63,p=0.004) (Figure 4). The
overall ΔIMCL CH3:CH2 significantly decreased during the
fast (both p<0.015; paired samples t-test). In each muscle, ΔIMCL CH3:CH2
did not correlate with any corresponding ΔEMCL measure.
Discussion
We have investigated the response of different fat stores to a 28 h fast and shown significant increases in CH2 IMCL in both the TA and SOL muscles, and significant decreases in both abdominal and leg subcutaneous fat. Increases in TA and SOL IMCL correlated with oleic and palmitic acid plasma FFA concentrations respectively, suggesting a differing uptake of fats between muscle groups during the fast. The compositional change of IMCL during the fast related inversely to the baseline composition, suggesting a selective efflux of unsaturated shorter chain IMCLs for preferential oxidation 1, which is in agreement with rat biopsy data 6, and supportive of the idea of IMCL as a flexible lipid store 7.Acknowledgements
AS is funded by the NIHR via an award to the Cambridge NIHR/WT Clinical Research Facility. We thank S. Nutland for participant recruitment, and acknowledge research grants from Addenbrooke’s Charitable Trust and the British Society for Pediatric Endocrinology and Diabetes.References
1. Price E.,Valencak T.G., 2012, In ‘Comparative Physiology of Fasting, Starvation, and Food Limitation’, 237-255
2. Halliwell K.J. et al., 1996, J. Lipid Res., 37, 1842-1848
3. Naressi A. et al., 2001, MAGMA, 12, 141-152
4. Vanhamme L. et al.,1997, J. Magn. Reson., 129, 35-43
5. Ren J. et al., 2010, MRM, 64, 662-671
6. Herzberg G.R., Farrell B., 2003, Nut. Res., 23, 205-213
7. Loher H. et al., 2016, Int. J. Mol. Sci., 17, 1554
Figures