2475
Disrupted topological organization of brain networks in early Parkinson’s disease (PD) subjects: Insights from Parkinson’s Progressive Markers Initiative (PPMI) dataset1Cleveland Clinic Lou Ruvo Center for Brain Health, Las Vegas, NV, United States
Synopsis
In vivo imaging that reliably captures the impact of the spreading pathology of Parkinson’s disease (PD), including its impact on both white and gray matter, remains elusive. In this study, we applied graph-theoretical techniques to multi-site diffusion-MRI data from a cohort of early PD-subjects in Parkinson’s Progressive Markers Initiative (PPMI) database. A disrupted topological brain organization in early PD-subjects was revealed with impaired network integration, segregation, global efficiency, and local efficiency. Furthermore, there was also
Introduction
The pathologic development of Parkinson’s disease (PD) appears to be related to the spread of abnormal synuclein in a largely caudal-rostral direction in the CNS1–3. In vivo imaging that captures the impact of this spreading pathology on both white and gray matter remains elusive4,5. Graph theoretical approaches have the ability to characterize connectivity at both global and local levels and can provide useful information about structural brain-network disorganization6, but this has not been validated in a multi-site cohort of early PD-subjects to the best of our knowledge. Hence, we applied graph theoretical approaches to diffusion-MRI data from Parkinson’s Progressive Markers Initiative (PPMI) database7 and showed topological differences in network properties which may in turn help differentiate PD from controls early in the disease.Methods
Subjects: Diffusion-MRI and T1-weighted MRI data from 69 (24 female) healthy controls (age: 60.61±10.61years, years of education (YOE): 15.61±3.07) and 151 (53 female) early PD-subjects (age: 61.03±9.23years, YOE: 15.14±2.95, total MDS-UPDRS: 30.81±13.03; disease duration: 7±7.14months) were derived from PPMI for this study. Imaging parameters are described in detail at http://www.ppmi-info.org/7. Only data from 3T Siemens scanners with the first visit were used to ensure uniformity of diffusion data. Network construction: Two different atlases, AAL (mainly cortical)8, and ATAG (subcortical)9 in MNI space, were used to generate 102 nodes (90 AAL-nodes and 12 ATAG-nodes) of the network. T1-weighted MNI152 brain was normalized to each subject’s native diffusion space and the resultant transformation matrix was applied to both the atlases to get the nodes in subject’s native space. Whole brain tractography was performed using diffusion toolkit (http://www.trackvis.org/dtk/)10. The nodes were expanded by four voxels11 and only those fiber-tracts that have ends in either nodes were retained. Fibers smaller than 6mm and internode connectivity with less than 10 fibers were filtered from any further analysis. Each internode connection (edge) was weighted by the product of the number of fibers and average FA of the fibers connecting the two nodes and normalized by the product of average length of the fibers and summation of volume of the two nodes. Graph-theoretical measures: Global(GE) and local efficiency(LE), pathlength(Lp), and clustering coefficient(Cp) were computed using GRETNA12 for each subject. Each measure was computed at various sparsity thresholds(5-40%, step=1%). Community assignment and nodal measures of efficiency, Lp, and Cp were extracted at a sparsity at which the network was fully connected for both groups13. Statistical analysis: Two-sample-t-test was conducted to see if any of the measures are significantly different between two-groups at any threshold.Results
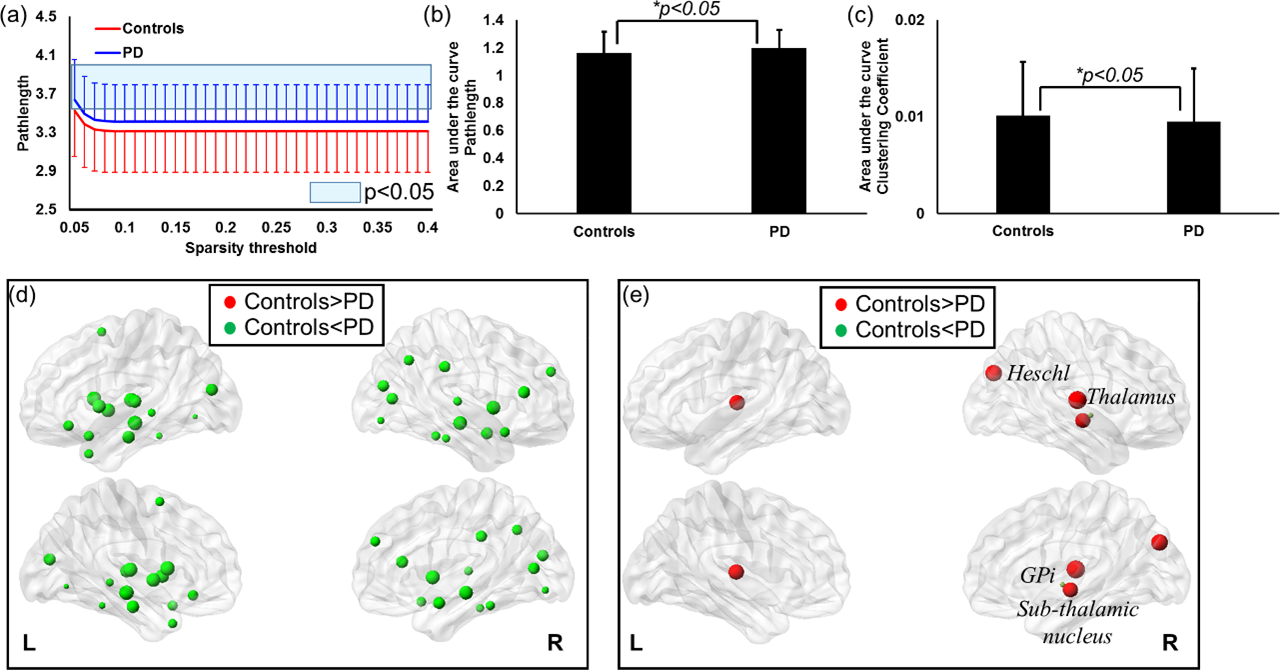
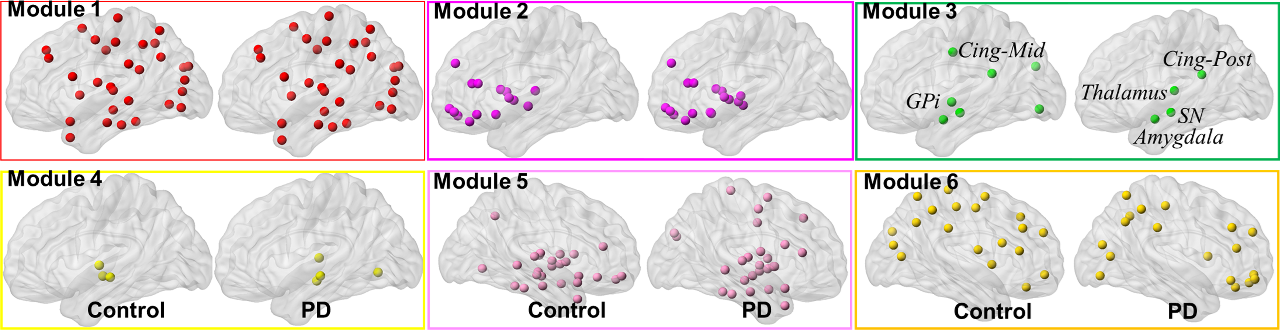
Most of the major cortical-subcortical fibers(Fig.1(a)) were retained (representative healthy control (top-row) and PD-subject (bottom-row)) as internode fibers, as expected. The mean connectivity matrix (Fig.1(c)) also does not reveal any qualitative differences between the groups. Whole brain analysis, however, revealed significantly(p<0.05) lower GE(Fig.2 (a) and (c)) and LE(Fig.2 (b) and (d)) in PD-subjects across all sparsity thresholds. Nodal comparisons at a sparsity of 10% revealed Globus-Pallidum-internal (GPi), striatum, and olfactory gyrus with significantly(p<0.05) higher LE in PD than controls and visualized in Fig.2(f). No nodes were found to have a higher GE in PD subjects (Fig.2(e)). Significantly longer Lp (reduced network integration)(p<0.05) was observed in PD at every sparsity threshold (Fig.3 (a) and (b)). No significant differences were obtained for Cp at any sparsity threshold but area under the curve revealed significantly (p<0.05) higher Cp (better network segregation) in controls (Fig.3(c)). Nodal analysis revealed no nodes with a longer Lp in controls and only GPi with higher Cp in PD (Fig.3(d)). Nodes with longer Lp in PD were located at middle temporal lobe, inferior frontal gyrus, and insula (Fig.3(e)). Six modules were found in both the groups and were not found to be significantly different between the groups (Fig.4). However, there was a differential assignment of few nodes to different modules between controls and PD.Discussion
PD-subjects revealed impaired network-integration and segregation as observed through significantly longer Lp, and lower Cp along with lower GE and LE as compared to controls. Importantly, nodes that exhibited significantly different network properties are implicated early in PD1,2. Furthermore, there was also rearrangement of nodes in different modules between the groups. Thus, without a priori modeling we detected organization in brain network known to be important in PD. Future studies will expand investigation of changes in these network properties and disease progression and severity, and evaluate any pattern shift in the modules as well.Conclusion
Graph theoretical study of early PD subjects revealed disrupted topological organization in PD subjects which can be exploited in understanding disease progression in PD subjects.Acknowledgements
The study was supported in parts by National Institute of General Medical Sciences (grant: P20GM109025) and the Elaine.P.Wynn and family foundation.References
1. Braak H, Del Tredici K, Rub U, de Vos RAI, Jansen Steur ENH, Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging 2003; 24: 197–211.
2. Braak H, Bohl JR, Muller CM, Rub U, de Vos RAI, Del Tredici K. Stanley Fahn Lecture 2005: The staging procedure for the inclusion body pathology associated with sporadic Parkinson’s disease reconsidered. Mov Disord 2006; 21: 2042–51.
3. Goedert M, Spillantini MG, Del Tredici K, Braak H. 100 years of Lewy pathology. Nat Rev Neurol 2013; 9: 13–24.
4. Hall JM, Ehgoetz Martens KA, Walton CC, et al. Diffusion alterations associated with Parkinson’s disease symptomatology: A review of the literature. Parkinsonism Relat Disord 2016; published online Sept. DOI:10.1016/j.parkreldis.2016.09.026.
5. Pyatigorskaya N, Gallea C, Garcia-Lorenzo D, Vidailhet M, Lehericy S. A review of the use of magnetic resonance imaging in Parkinson’s disease. Ther Adv Neurol Disord 2014; 7: 206–20.
6. Bullmore E, Sporns O. Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci 2009; 10: 186–98.
7. www.ppmi-info.org.
8. Tzourio-Mazoyer N, Landeau B, Papathanassiou D, et al. Automated anatomical labeling of activations in SPM using a macroscopic anatomical parcellation of the MNI MRI single-subject brain. Neuroimage 2002; 15: 273–89.
9. Keuken MC, Bazin P-L, Crown L, et al. Quantifying inter-individual anatomical variability in the subcortex using 7 T structural MRI. Neuroimage 2014; 94: 40–6.
10. Wang R, Wedeen VJ. TrackVis.org. In: Proc Intl Soc Mag Reson Med. 2007: 3720.
11. Jeon T, Mishra V, Huang H. Effects of cortical regions of interests on tractography and brain connectivity quantification. In: Proc Intl Soc Mag Reson Med. 2016: 2063.
12. Wang J, Wang X, Xia M, Liao X, Evans A, He Y. GRETNA: a graph theoretical network analysis toolbox for imaging connectomics. Front Hum Neurosci 2015; 9: 386.
13. He Y, Chen Z, Evans A. Structural insights into aberrant topological patterns of large-scale cortical networks in Alzheimer’s disease. J Neurosci 2008; 28: 4756–66.
Figures