1691
Connectivity of the Default Mode Network – Prior and post an oddball paradigm1Institute of Neuroscience and Medicine 4 (INM4), Forschungszentrum Juelich GmbH, Juelich, Germany, 2Department of Psychiatry, Psychotherapy and Psychosomatics, RWTH Aachen University, Aachen, Germany, 3JARA – BRAIN – Translational Medicine, Juelich, Germany, 4TRIMAGE-consortium, Juelich, Germany, 5Department of Neurology, RWTH Aachen University, Aachen, Germany, 6Department of Electrical and Computer Systems Engineering, and Monash Biomedical Imaging, School of Psychological Sciences, Monash University, Melbourne, Australia
Synopsis
The DMN is shown to be highly active during resting state and is rapidly deactivated during externally directed tasks. However, it is unclear how demanding the task needs to be to switch the DMN out of the resting state mode. Aiming to answer this question we chose a visual oddball paradigm and assessed the functional connectivity of the DMN during resting state in a rest-task-rest design. The oddball paradigm demands very little cognitive resources and therefore we hypothesize that the DMN will not be altered in its connectivity pattern during post task resting state.
Aim
The default mode network (DMN) is thought to characterize basal neural activity 1,2 and has been linked to self-referential thought, introspection and integration of cognitive, and emotional processing 3. It has been shown to be highly active during resting state and is rapidly deactivated during externally directed tasks 4. However, it is unclear how demanding the task needs to be to switch the DMN out of the resting state mode. Aiming to answer this question we chose a visual oddball paradigm6 and assessed the functional connectivity of the DMN in a rest 1 - task - rest 2 design. This oddball paradigm demands very little cognitive resources and therefore we hypothesize that the DMN will not be altered in its connectivity pattern.Methods
Resting state fMRI data were recorded in 22 male healthy volunteers (mean age=24 years, SD=4,3 years) prior to and post the P300 paradigm, in a 3T MR system (Trio, Siemens, Germany) using an EPI protocol (TR 3000msec, echo time 30ms, 6 minutes, eyes closed). The P300 paradigm involved a visual oddball task using yellow circles as frequent stimuli, blue ones as target stimuli; 3 runs á 200 trials: 160 frequent stimuli: passive viewing, button press for targets, counting targets8. Structural MRI images were also acquired using the MPRAGE protocol (TR 2250msec, echo time 3.03msec, FA 9°, 176 slices, FOV 256 x 256 mm, voxel size 1mm isotropic). A comparative connectivity analysis of DMN-PCC with the whole brain pre and post task was performed, after preprocessing (slice time correction, normalization, smoothing, realignment, motion correction, de-noising using CompCor11, temporal band pass filtering 0.008-0.09Hz), using the SPM based toolbox CONN v15.h (http://www.nitrc.org/projects/conn). The analyses included ROI-ROI functional (correlation) and effective (regression) connectivities. ROI Atlases used for this purpose were the whole brain atlas (Cortical ROIs from Harvard-Oxford Maximum Likelihood atlas, Cerebellar parcellation from AAL atlas) and default mode network atlas4 both included with CONN. Additionally, graph theory was applied to thresholded connectivity matrices to investigate changes in graph topology.Results
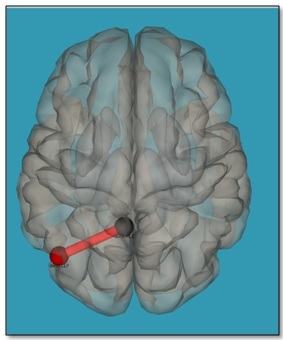
Connectivity analysis showed no difference in PCC-ROI connectivity pre and post task (p-FDR corrected <0.05). Additionally no significant changes in graph topology were observed for all thresholds from 0.25 to 0.75 (two sided, p-FDR corrected < 0.05). However, a trend for a slightly increased effective connectivity between PCC and the left lateral parietal (LLP) region was observed on liberalizing the significance threshold (p-uncorrected < 0.001) (Figure 1). This was also checked by data splitting with 5 combinations of 16 subjects.Discussion
The visual oddball task did not change the connectivity of PCC, a main hub of the DMN, to other brain areas. However, when lowering the threshold to an uncorrected level (p<0.001) a trace of the given task is observed: an increased effective connectivity between the PCC and the left lateral parietal cortex. These results are in line with our hypothesis that the P300 paradigm requires the allocation of attention to the stimuli for the discrimination of target and non-target stimuli, but this allocation of attention lies below the threshold for switching the DMN out of resting state. This might be discussed as an ongoing filtering of the "outer world" at a low level as ongoing basal neural activity while the brain is engaged in a "self-referential mode" during resting state.Conclusion
Even during resting state the brain holds up a basic filtering of the "outer world" without strong alterations of the DMN connectivity patterns.Acknowledgements
No acknowledgement found.References
1. Raichle, M. E. & Snyder, A. Z. A default mode of brain function: A brief history of an evolving idea. (2007). doi:10.1016/j.neuroimage.2007.02.041
2. Snyder, A. Z. & Raichle, M. E. A brief history of the resting state: The Washington University perspective. Neuroimage 62, 902–910 (2012).
3. Greicius, M. D., Krasnow, B., Reiss, A. L. & Menon, V. Functional connectivity in the resting brain: A network analysis of the default mode hypothesis. 4. Fox, M. D. et al. The human brain is intrinsically organized into dynamic, anticorrelated functional networks. (2005).
5. Vogt, B. A., Finch, D. M. & Olson, C. R. Functional heterogeneity in cingulate cortex: the anterior executive and posterior evaluative regions. Cereb. Cortex 2, 435–43
6. Warbrick, T., Reske, M. & Shah, N. J. Do EEG paradigms work in fMRI? Varying task demands in the visual oddball paradigm: Implications for task design and results interpretation. Neuroimage 77, 177–185 (2013).
7. Warbrick, T. et al. Single-trial P3 amplitude and latency informed event-related fMRI models yield different BOLD response patterns to a target detection task. Neuroimage 47, 1532–1544 (2009).
8. Warbrick, T., Arrubla, J., Boers, F., Neuner, I. & Shah, N. J. Attention to Detail: Why Considering Task Demands Is Essential for Single-Trial Analysis of BOLD Correlates of the Visual P1 and N1. J. Cogn. Neurosci. 26, 529–542 (2014).
9. Jenkinson, M., Bannister, P., Brady, M. & Smith, S. Improved Optimization for the Robust and Accurate Linear Registration and Motion Correction of Brain Images. Neuroimage 17, 825–841 (2002).
10. Smith, S. M. Fast robust automated brain extraction. Hum. Brain Mapp. 17, 143–55 (2002).
11. Whitfield-Gabrieli, S. & Nieto-Castanon, A. Conn: A Functional Connectivity Toolbox for Correlated and Anticorrelated Brain Networks. doi:10.1089/brain.2012.0073
12. Smith, S. M. et al. Correspondence of the brain’s functional architecture during activation and rest.
Figures