1690
Spatio-temporal extent of spontaneous and sensory evoked neural network activity.1Institute for Microscopic Anatomy and Neurobiology, Johannes Gutenberg-University Mainz, Mainz, Germany, 2Department of Clinical Radiology, University of Münster, Nordrhein-Westfalen, Germany
Synopsis
Preclinical functional Magnetic Resonance Imaging in rodents has become a vital research tool for observing functional network connectivity at rest or during sensory stimulation. There is an ongoing debate about the differential effects of anesthesia on cortical connectivity, since functional connectivity networks fluctuate depending on global brain state. We analyzed the spontaneous and sensory evoked BOLD signal in rats during different network states induced by Isoflurane and Medetomidine anesthesia, finding them to lead to fundamentally different cortical connectivity. These results are crucial for interpreting rodent studies in the framework of translational resting state research including awake human data.
PURPOSE
Preclinical functional Magnetic Resonance Imaging (fMRI) in rodents has become a vital research tool for observing functional network connectivity at rest or during sensory stimulation in health and disease. In humans, resting state studies are performed in awake individuals. In contrast, in animals most of these studies are carried out under anesthesia, which can dramatically modify global cortical connectivity[1]. Default thalamocortical network activity underlying the BOLD signal can be altered by anesthesia and determines BOLD activity at rest as well as during responses to sensory stimulation[2]. Neurophysiological studies employing cellular readouts show that such network activity - reflected by default firing modes as well as response properties upon sensory stimulation of local neural networks - are dramatically altered by behavioral state[3], sleep or wakefulness[4], as well as anesthetic regimen[5-7]. Consequently, there is still a debate on differential effects of anesthesia on cortical connectivity during resting state or sensory stimulation, since functional connectivity networks are fluctuating depending on brain states. Optic-fiber based calcium recordings reflect reflect suprathreshold neuronal activity (spiking) on population level and allow the monitoring of neuronal firing modes of a local neuronal population. Here, by comparing BOLD functional connectivity and optic-fiber based calcium recordings, we demonstrate that different brain states are being evoked by the two distinct anesthetic regimens, which govern functional connectivity at rest and during sensory stimulation.METHODS
For optic-fiber based calcium recordings, anesthetized rats were placed in a stereotactic frame and a craniotomy was conducted at the level of primary somatosensory cortex, Oregon Green 488 BAPTA-1 AM (OGB-1) was injected and an optical fiber was implanted. A custom-made optical laser setup was used to excite the calcium dye and to record changes in fluorescence. For electric stimulation, two needle electrodes were inserted into the forepaw. Functional MRI was performed in a 9.4 T Biospec (Bruker). BOLD and ICA analysis were performed with SPM8. For the sensory stimulation condition we used the activated somatosensory cortex as seed to analyze correlations of the BOLD signal of this region with others previously defined. Functional connectivity analysis was performed with a Matlab script using 96 atlas-based cortical regions of the rat brain[8] during spontaneous activity as well as during sensory stimulation.RESULTS
Employing optic-fiber calcium recordings alongside fMRI measurements in rats, we showed rhythmic slow calcium waves present in the cortex and thalamus under isoflurane anesthesia (1.3-1.8%) (Figure 1 A, B). Using onsets and durations of these events as regressor, an event-related fMRI analysis revealed pancortical BOLD activation, speaking for synchronous cortical activity related to slow calcium waves (Figure 1 C). In contrast, in a rather sedative state induced by Medetomidine, the defined on- and off-periods of neuronal firing as in slow wave state are absent (Figure 1, D). Sensory responses are fundamentally different from the ones during slow wave network activity, as revealed by calcium recordings in the two states (Figure 1 D). Independent Component (ICA) and correlation analysis also reveal a pancortical spread of BOLD activation during slow wave activity (Figure 1 E). For Medetomidine sedation ICA and correlation analysis do not show a defined widespread BOLD activation but a rather patterned signal when analyzing activity unspecific for sensory stimulation (Figure 1 F) and a defined, region-specific activity when using the somatosensory cortex as a seed region (Figure 1 G).DISCUSSION
As local cortical calcium recordings in these states reflect profoundly different firing modes, we hypothesized that they may be linked to different cortical functional connectivity states, reflecting different global brain states. These results give inside about the actual underlying cellular activity which the BOLD signal alone cannot reveal, and could explain variations of sensory-evoked BOLD responses under different anesthetics leading to different brain states.CONCLUSION
Here, we unambiguously classified brain states by using a secondary readout such as calcium recordings, ideally in parallel to fMRI scans. The different types of network activation revealed in this study strongly suggest, that the effect of brain states should be taken into account upon interpreting anesthesia based rodent studies in the framework of translational resting state research including awake human data.Acknowledgements
No acknowledgement found.References
1. Liu, X., et al., The change of functional connectivity specificity in rats under various anesthesia levels and its neural origin. Brain Topogr, 2013. 26(3): p. 363-77.
2. Tu, Y., et al., Altered thalamocortical functional connectivity by propofol anesthesia in rats. Pharmacology, 2011. 88(5-6): p. 322-6.
3. McGinley, M.J., et al., Waking State: Rapid Variations Modulate Neural and Behavioral Responses. Neuron, 2015. 87(6): p. 1143-61.
4. Steriade, M., et al., The slow (< 1 Hz) oscillation in reticular thalamic and thalamocortical neurons: scenario of sleep rhythm generation in interacting thalamic and neocortical networks. J Neurosci, 1993. 13(8): p. 3284-99.
5. Constantinople, C.M. and R.M. Bruno, Effects and mechanisms of wakefulness on local cortical networks. Neuron, 2011. 69(6): p. 1061-8.
6. Schmid, M.C. and A. Maier, To see or not to see--thalamo-cortical networks during blindsight and perceptual suppression. Prog Neurobiol, 2015. 126: p. 36-48.
7. Stroh, A., et al., Making waves: initiation and propagation of corticothalamic Ca2+ waves in vivo. Neuron, 2013. 77(6): p. 1136-50.
8. Valdes-Hernandez, P.A., et al., An in vivo MRI Template Set for Morphometry, Tissue Segmentation, and fMRI Localization in Rats. Front Neuroinform, 2011. 5:p. 26.
Figures
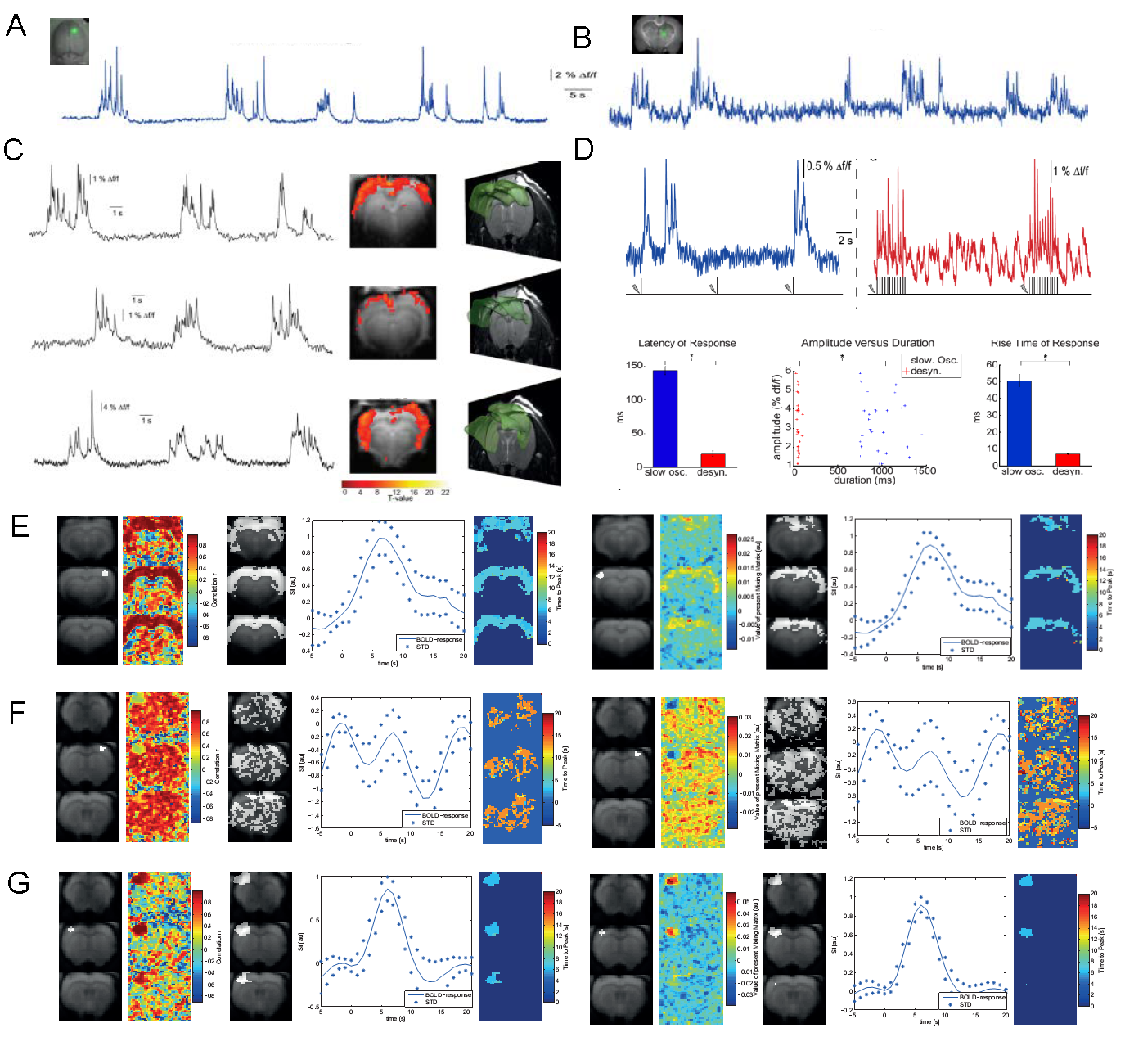
A. OGB-1 in somatosensory cortex and fluorescent changes reflecting neural network activity in isoflurane-induced slow wave state.
B. Same than A in thalamus.
C. Slow calcium waves simultaneously recorded to BOLD-fMRI. Onsets and durations of waves reveal pancortical BOLD activity within an event-related fMRI analysis.
D. Slow wave activity (SWA) induced by isoflurane and persistent activity induced by Medetomidine leads to different calcium signal responses upon sensory stimulation.
E. Correlation (left) and ICA (right) reveal pancortical BOLD activation during SWA.
F. Same analysis reveal unspecific activity under Medetomidine sedation.
G. During sensory stimulation Medetomidine sedation leads to region-specific BOLD actiivation.