5027
Effect of Caffeine on Blood-Brain Barrier Water Permeability Measured with Intrinsic Diffusivity Encoding of Arterial Labeled Spins (IDEALS)1Biomedical Engineering, Stony Brook University, Stony Brook, NY, United States, 2Radiology, Stony Brook University, Stony Brook, NY, United States
Synopsis
Caffeine is a commonly used neurostimulator that also produces cerebral vasoconstriction by antagonizing adenosine receptors. Previous studies demonstrate that acute caffeine intake leads to significant cerebral blood flow (CBF) reduction but have not investigated the effect on blood-brain barrier (BBB) water permeability. Here we provide an initial investigation into the effect of caffeine on BBB water permeability parameters, water extraction fraction (Ew) and permeability surface area product (PSw), using the recently developed Intrinsic Diffusivity Encoding of Arterial Labeled Spins (IDEALS). Significant reductions in CBF, Ew, and PSw were observed after administration of 200 mg caffeine.
Introduction
Blood-brain barrier (BBB) disruption has been linked to multiple central nervous system diseases and psychiatric disorders including cancer, multiple sclerosis, stroke, and mood, behavior and cognitive dysfunctions1. Caffeine is a commonly used neurostimulator that also produces cerebral vasoconstriction by antagonizing adenosine receptors2. Although the underlying mechanism are not fully understood, caffeine has been shown in human and animal studies to protect against Alzheimer’s and Parkinson’s diseases3. Acute caffeine intake leads to significant cerebral blood flow (CBF) reduction4. Here we provide an initial investigation into the effect of caffeine on BBB water permeability parameters, water extraction fraction (Ew) and permeability surface area product (PSw), using the recently developed Intrinsic Diffusivity Encoding of Arterial Labeled Spins (IDEALS) technique5,6.Methods
Four healthy participants were recruited with IRB approval and informed consent. All studies were performed on a Siemens 3T Prisma MRI with 64-channel head/neck coil. Subjects were instructed to continue normal caffeine consumption but to abstain from caffeine intake for ~6 hours prior to the exam. Subjects were imaged at baseline, then taken out of the scanner, but remained on the table, and ingested a 200 mg caffeine pill. After ~10 minutes of rest, subjects were repositioned and imaged. In the IDEALS paradigm, intravascular and extravascular ASL water are separated by their different diffusion sensitivities at two segmentation factors in 3D-GRASE acquisition7. The MRI parameters were: TR/TE/Label Time 4500/16/1600 ms, FA=120°, matrix of 64×64×32, FOV of 256×256×128 mm3, iPAT2. Two segmentation schemes (4PAR×2PE and 1PAR×2PE) and two PLDs (1000 and 2000 ms) were used for a total of 4 sets of ASL images with total acquisition time ~15 min. High resolution MPRAGE images were acquired for tissue segmentation. The arterial transit time (ATT) was estimated from the 1000 ms PLD data. CBF, Ew, and PSw were subsequently estimated from the 2000 ms PLD data. Two-tailed paired t-tests were used to test for significant differences in gray matter (GM) and white matter (WM) before and after caffeine challenge; p < 0.05 was considered significant.Results
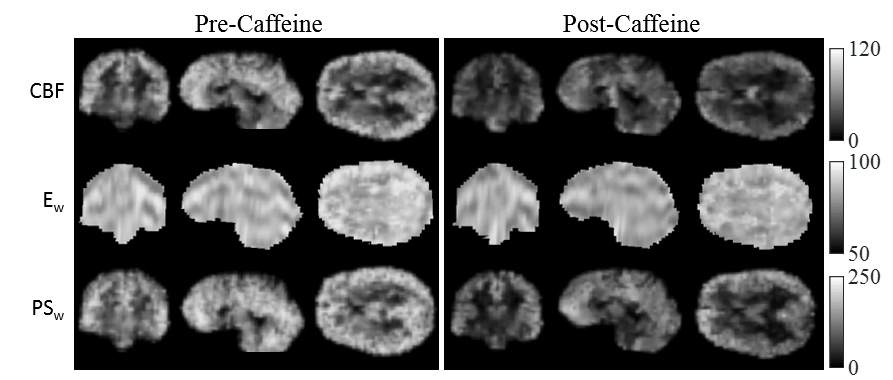
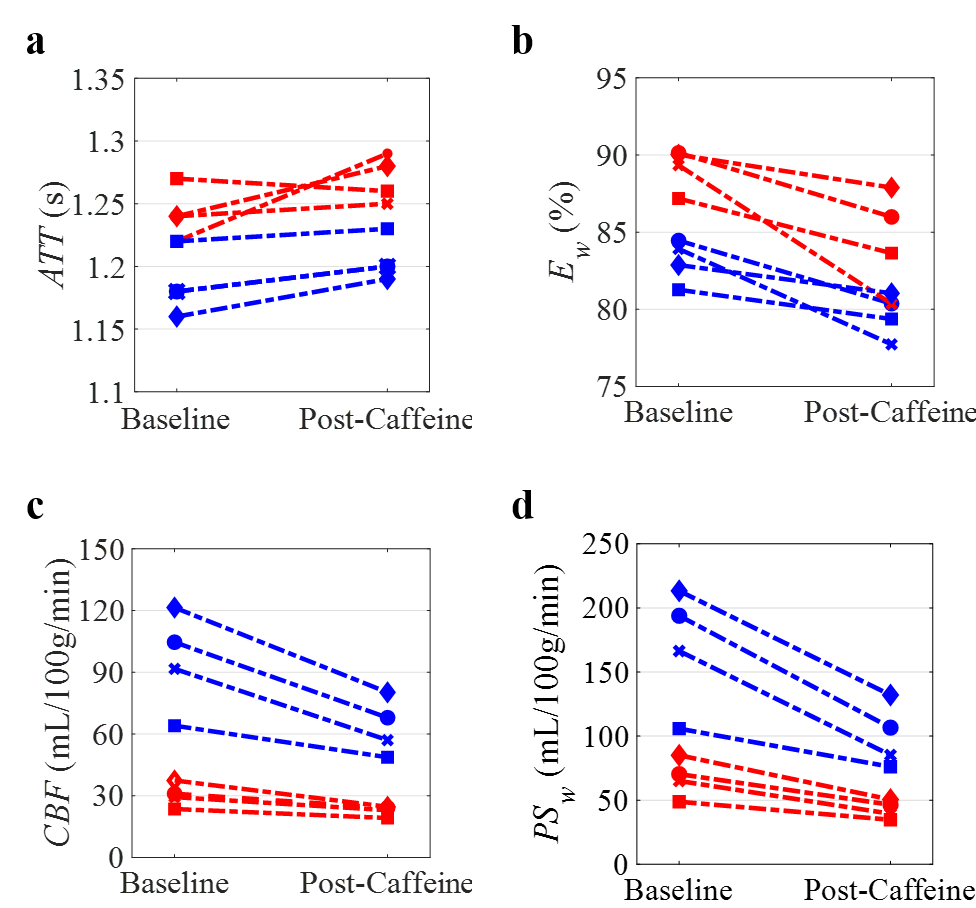
Figure 1 displays the derived whole-brain BBB permeability maps from a healthy participant before and after caffeine challenge. Figure 2 shows the ladder plots of CBF, ATT, Ew, and PSw for the 4 subjects imaged before and after caffeine challenge. In this study, acute caffeine intake significantly reduced CBF in both GM (95.5 ± 24.2 vs. 63.5 ± 13.7ml/100g/min) and WM (30.3 ± 5.7 vs. 22.8 ± 2.5 ml/100g/min). Accordingly, the estimated ATT increased ~2% from baseline in both GM (1.18 ± 0.03 vs. 1.21 ± 0.01 ms; p < 0.05) and WM (1.24 ± 0.02 vs. 1.26 ± 0.02 ms; p = 0.27). In both GM and WM, significant reductions to both Ew (83.1% ± 1.4% vs. 79.6 ± 1.4% in GM; 89.2% ± 1.4% vs. 84.5% ± 3.2% in WM) and PSw (169.9 ± 46.8 vs. 100.0 ± 24.9mL/100g/min in GM; 67.3 ± 15.0 vs. 42.8 ± 7.0mL/100g/min in WM) were observed.Discussion
Caffeine affects neurotransmitter release and neuronal firing rates via action of adenosine receptors. A recent study in a rat chronic sleep restriction model reported decreased BBB permeability to dextrans and Evans blue after administration of an A2A adenosine receptor antagonist8. Although there is no existing literature on the role of adenosine receptors in BBB water permeability, its activation in the collecting duct of kidneys is known to reduce water permeability and transport9. Our preliminary study is the first to assess the effect of caffeine on BBB water permeability in human subjects in-vivo. The observed reductions in Ew and PSw after caffeine intake may represent the combined effect from reduced capillary water permeability (through adenosine A1 receptor activation), reduced microvascular surface area (from binding to adenosine A2A and A2B receptors on vascular smooth muscle), and the inhibition of ATP-dependent K+ channels (water co-transporter)10-13. Therefore, the BBB protection effect from chronic caffeine-use may come from increased adenosine receptor density in cerebral endothelial cells14.Conclusion
Changes to BBB water permeability in response to caffeine challenge were measured using the IDEALS approach. Significant decreases to CBF, Ew, and PSw were observed after administration of 200 mg of caffeine.Acknowledgements
No acknowledgement found.References
1. Hawkins BT, Davis TP. The blood-brain barrier/neurovascular unit in health and disease. Pharmacological reviews. 2005;57(2):173-185.
2. Fredholm BB. Adenosine, adenosine receptors and the actions of caffeine. Pharmacology & toxicology. 1995;76(2):93-101.
3. Chen X, Ghribi O, Geiger JD. Caffeine protects against disruptions of the blood-brain barrier in animal models of Alzheimer's and Parkinson's diseases. Journal of Alzheimer's Disease. 2010;20(s1):S127-S141.
4. Addicott MA, Yang LL, Peiffer AM, et al. The effect of daily caffeine use on cerebral blood flow: How much caffeine can we tolerate? Human brain mapping. 2009;30(10):3102-3114.
5. He X, Wengler K, Duong T, Schweitzer M. 3D MRI Mapping of Whole-Brain Water Permeability with Intrinsic Diffusivity Encoding of Arterial Labeled Spins (IDEALS). In Proceedings of the 27th Annual Meeting of ISMRM, Paris. 2018;0180.
6. Wengler K, Ha J, Coyle P, Schweitzer M, Duong T, He X. Blood Brain Barrier Water Permeability in Non-Enhancing Multiple Sclerosis Lesion with Intrinsic Diffusivity Encoding of Arterial Labeled Spins (IDEALS). In Proceedings of the 27th Annual Meeting of ISMRM, Paris. 2018;4921.
7. He X, Wengler K, Schweitzer ME. Diffusion sensitivity of 3D‐GRASE in arterial spin labeling perfusion. Magnetic resonance in medicine. 2018;80(2):736-747.
8. Hurtado-Alvarado G, Domínguez-Salazar E, Velázquez-Moctezuma J, Gómez-González B. A2A adenosine receptor antagonism reverts the blood-brain barrier dysfunction induced by sleep restriction. PloS one. 2016;11(11):e0167236.
9. Rieg T, Vallon V. ATP and adenosine in the local regulation of water transport and homeostasis by the kidney. American Journal of Physiology-Regulatory, Integrative and Comparative Physiology. 2009;296(2):R419-R427.
10. Lee J, Ha JH, Kim S, Oh Y, Kim SW. Caffeine decreases the expression of Na+/K+‐ATPase and the type 3 Na+/H+ exchanger in rat kidney. Clinical and experimental pharmacology and physiology. 2002;29(7):559-563. 1
1. Bai R, Springer Jr CS, Plenz D, Basser PJ. Fast, Na+/K+ pump driven, steady‐state transcytolemmal water exchange in neuronal tissue: A study of rat brain cortical cultures. Magnetic resonance in medicine. 2018;79(6):3207-3217.
12. Bai R, Springer Jr CS, Plenz D, Basser PJ. Brain active transmembrane water cycling measured by MR is associated with neuronal activity. Magnetic resonance in medicine. 2018.
13. Kleppisch T, Nelson MT. Adenosine activates ATP-sensitive potassium channels in arterial myocytes via A2 receptors and cAMP-dependent protein kinase. Proceedings of the National Academy of Sciences. 1995;92(26):12441-12445.
14. Shi D, Nikodijević O, Jacobson KA, Daly JW. Chronic caffeine alters the density of adenosine, adrenergic, cholinergic, GABA, and serotonin receptors and calcium channels in mouse brain. Cellular and molecular neurobiology. 1993;13(3):247-261.
Figures