4915
Assessment of two T1D components within myelinated tissue with ihMT MRI1Aix Marseille Univ, CNRS, CRMBM UMR 7339, Marseille, France, 2Aix Marseille Univ, CNRS, ICR UMR 7273, Marseille, France, 3CBMN UMR 5248, CNRS University of Bordeaux, Pessac, France, 4Department of Radiology, Division of MR Research, Beth Israel Deaconess Medical Center, Harvard Medical School, Boston, MA, United States
Synopsis
T1D, the relaxation time of the dipolar order, is a probe for membrane dynamics and organization that could be used to assess myelin integrity. A single-component T1D model associated with a modified ihMT sequence had been proposed for in vivo evaluation of T1D with MRI. However, experiments and simulations revealed that myelinated tissues exhibit multiple T1D components. A bi-component T1D model is proposed and validated. Fits in a rat spinal cord yield two T1Ds of about 10 ms and 400 μs. The results suggest that myelin has a dynamically heterogeneous organization.
Motivation: T1D as a probe of membrane molecular dynamics.
Nuclear spin relaxation is characterized by relaxation times (e.g. T1 and T2) which are sensitive to different ranges of motional frequencies. So nuclear spin relaxation can in principle deliver quantitative information about membrane molecular dynamics and organization1-2, such as correlation times and activation energy to describe motions.
T1D, the relaxation time of the dipolar order, can provide new information about membrane dynamics. The dipolar order is built due to the local fields of dipolar-coupled spins3, which are much weaker than the external magnetic fields. Thus, as compared to T1, T1D is more sensitive to slower motional processes, such as membrane collective motions4-5 (Fig.1A).
Collective motions in a membrane are hydrodynamical deformations, such as bending, stretching and compression. They could be probed to quantify membrane fluidity and elasticity. This is interesting for myelin, a membrane that is particularly important to study6, since assessing myelin fluidity may yield further understanding of demyelinating diseases, such as multiple sclerosis7.
IhMT, inhomogeneous magnetization transfer8, is a T1D-weighted MRI technique, whose measurement sequence can be tailored to enable T1D measurements9 (Fig.1B).
Background: limitations of the single-component T1D model.
The ihMT model presented in [8] considers only one dipolar reservoir (Fig.2A), and therefore a single T1D value.
This single-component T1D model was used to fit T1D of a rat spinal cord ex-vivo for different irradiation RF powers (B1RMS). The experimental parameters are listed in Table 1. The results (Fig.2B-C) show that the apparent T1D values in both white (WM) and gray matter (GM) decrease as B1RMS is increased.
This dependence on power could be explained by the existence of multiple T1D components (long and short T1D) in spinal cord. If we assume that, in lipids, there are several components that contribute to ihMT, each one with different molecular dynamics, each component would be associated with a different T1D10. At lower power, only the long T1Ds are revealed and contribute to ihMTR signal11. However, since the single-component T1D model only reports one apparent T1D, which is expected to represent a weighted average of all T1D components, it tends to reduce T1D values when power is increased as short T1D components are revealed.
This phenomenon was further confirmed by simulations. By using a bi-component T1D model, with two dipolar reservoirs (Fig.3A), noisy synthetic ihMTR data (Fig.3B) were generated with the values: T1D1=4 ms, T1D2=0.4 ms and fD=0.4, where fD represents the fraction of the semisolid pool associated with T1D1. The fraction associated with T1D2 is (1-fD).
Rician noise, with standard deviation set to 0.0001, was added 1000 times and the synthetic data were subsequently fitted with the single-T1D model (Fig. 2A). These steps were repeated for different B1RMS.
The same behavior as for the experimental data was observed: the apparent T1D decreases with B1RMS (Fig.3C). This suggests that the bi-component T1D model should be more appropriate to fit experimental data.
Proposed model: fitting two T1Ds.
To include a second dipolar reservoir in the ihMT model, we have applied the matrix formalism to the ihMT theory12 and solved the differential equations at the steady-state by using the matrix exponential solution13. The mathematical formalism of this model is detailed in the Appendix.
The same spinal cord experimental data were fitted again with the proposed bi-component T1D model (Fig.4). Long T1D (T1D1) in the order of 10 ms and short T1D (T1D2) in the order of 400 μs were found. In WM, the fraction of short T1D, (1-fD), increases with B1RMS, suggesting that short T1D components are revealed. Likewise, the short T1D component (T1D2) maps are revealed as B1RMS is increased, which is consistent with the theory11.
The increasing trend of T1D1 and T1D2 with power for B1RMS>5.8uT will require further investigations. At this stage, care must be taken when interpreting these results biophysically.
Conclusions: myelinated tissues contain multiple T1D components.
The existence of multiple T1D components in rat spinal cord is suggested by both experimental and simulated data. The proposed bi-component T1D model allowed measuring, in spinal cord, long and short T1D (in the order of 10 ms and 400 μs, respectively), the latter being revealed under strong RF power irradiation. Overall, the hypothesis of multi-component T1D in myelin is reasonable because myelin has a complex and dynamically heterogeneous organization.
Although a theory that would link the actual T1D values to membrane properties, such as fluidity and elasticity, is still missing, this work is an important step for assessing myelin integrity in vivo.
Appendix: the mathematical formalism of the proposed model.
Proposed model:
\begin{equation*}\dot{M}=AM+B\end{equation*}
where M is the magnetization of each reservoir (Fig.3A):
\begin{gather*}M=\begin{bmatrix}M_{ZA}\\M_{ZB1}\\\beta_{1}\\M_{ZB2}\\\beta_{2}\end{bmatrix}\end{gather*}
and A and B are:
\begin{gather*}A=\begin{bmatrix}-\left(R_{1A}+RM^{B}_{0}+R_{RFA}\right)&RM^A_0&0&RM^A_0&0\\Rf_DM^B_0&-\left(R_{1B}+RM^A_0+R_{RFB}\right)&R_{RFB}2\pi\Delta&0&0\\0&R_{RFB}\frac{2\pi\Delta}{D^2}&-\left[\frac{1}{T_{1D1}}+R_{RFB}\left(\frac{2\pi\Delta}{D}\right)^2\right]&0&0\\R(1-f_D)M^B_0&0&0&-\left(R_{1B}+RM^A_0+R_{RFB}\right)&R_{RFB}2\pi\Delta\\0&0&0&R_{RFB}\frac{2\pi\Delta}{D^2}&-\left[\frac{1}{T_{1D2}}+R_{RFB}\left(\frac{2\pi\Delta}{D}\right)^2\right]\\\end{bmatrix}\end{gather*}
and
\begin{gather*}B=\begin{bmatrix}R_{1A}M^A_0\\R_{1B}f_DM^B_0\\0\\R_{1B}(1-f_D)M^B_0\\0\end{bmatrix}.\end{gather*}
We want to know the values of M after a rectangular pulse13:
\begin{equation*}M_{t _1}= e^{At_1}M_{t=0}+A^{-1}\left(e^{At_1}-I\right)B.\end{equation*}
Then, we can calculate $$$ihMTR(Δt)$$$ at steady-state corresponding to all sequence variants shown in Fig.1B as:
\begin{equation*}ihMTR(Δt)=2\frac{M^+-M^{+-}(Δt)}{M_0}.\end{equation*}
Acknowledgements
This project has received funding from the European Union’s Horizon 2020 research and innovation programme under the Marie Skłodowska-Curie grant agreement No713750. Also, it has been carried out with the financial support of the Regional Council of Provence-Alpes-Côte d’Azur and with the financial support of the A*MIDEX (n° ANR- 11-IDEX-0001-02), funded by the Investissements d'Avenir project funded by the French Government, managed by the French National Research Agency (ANR).References
[1] Dufourc EJ, Mayer C, Stohrer J, Althoff G, Kothe G. Dynamics of phosphate head groups in biomembranes. Comprehensive analysis using phosphorus-31 nuclear magnetic resonance lineshape and relaxation time measurements. Biophys J. 1992;61(1):42-57.
[2] Trivikram R. Molugu, Soohyun Lee, and Michael F. Brown. Concepts and Methods of Solid-State NMR Spectroscopy Applied to Biomembranes. Chemical Reviews 2017 117 (19), 12087-12132. doi: 10.1021/acs.chemrev.6b00619
[3] Slichter, Charles P. Principles of Magnetic Resonance. Harper & Row Publishers, New York, 1963.
[4] R. Gaspar, E.R. Andrew, D.J. Bryant, and E.M. Cashell. Dipolar relaxation and slow molecular motions in solid proteins. Chemical Physics Letters, 86(4):327 – 330, 1982.
[5] O. Mensio, R.C. Zamar, F. Casanova, D.J. Pusiol, R.Y. Dong. Intramolecular character of the intrapair dipolar order relaxation in the methyl deuterated nematic para-azoxyanisole. Chemical Physics Letters 356 (2002) 457–461.
[6] Cornelia Laule, Irene M. Vavasour, Shannon H. Kolind, David K.B. Li, Tony L. Traboulsee, G.R. Wayne Moore, and Alex L. MacKay. Magnetic resonance imaging of myelin. Neurotherapeutics, 4(3):460 – 484, 2007. Advances in Neuroimaging/Neuroethics.
[7] Benjamin Ohler, Karlheinz Graf, Richard Bragg, Travis Lemons, Robert Coe, Claude Genain, Jacob Israelachvili, Cynthia Husted. Role of lipid interactions in autoimmune demyelination. Biochimica et Biophysica Acta 1688 (2004) 10–17
[8] G. Varma, O.M. Girard, V.H. Prevost, A.K. Grant, G. Duhamel, and D.C. Alsop. Interpretation of magnetization transfer from inhomogeneously broadened lines (ihMT) in tissues as a dipolar order effect within motion restricted molecules. Journal of Magnetic Resonance 260 (2015) 67–76.
[9] Gopal Varma, Olivier M. Girard, Valentin H. Prevost, Aaron K. Grant, Guillaume Duhamel, and David C. Alsop. In vivo measurement of a new source of contrast, the dipolar relaxation time, T1D, using a modified inhomogeneous magnetization transfer (ihMT) sequence. Magnetic Resonance in Medicine, 2017 Oct; 78(4): 1362-1372.
[10] Swanson, S. D., Malyarenko, D. I., Fabiilli, M. L., Welsh, R. C., Nielsen, J. and Srinivasan, A. (2017), Molecular, dynamic, and structural origin of inhomogeneous magnetization transfer in lipid membranes. Magn. Reson. Med., 77: 1318-1328. doi:10.1002/mrm.26210
[11] Alan P. Manning, Kimberley L. Chang, Alex L. MacKay, Carl A. Michal. The physical mechanism of ‘‘inhomogeneous” magnetization transfer MRI. Journal of Magnetic Resonance 274 (2017) 125–136.
[12] G. Varma, O.M. Girard, V.H. Prevost, A.K. Grant, G. Duhamel, and D.C. Alsop. Interpretation of magnetization transfer from inhomogeneously broadened lines (ihMT) in tissues as a dipolar order effect within motion restricted molecules. Journal of Magnetic Resonance 260 (2015) 67–76.
[13] Portnoy, S. and Stanisz, G. J. (2007), Modeling pulsed magnetization transfer. Magn. Reson. Med., 58: 144-155. doi:10.1002/mrm.21244
Figures
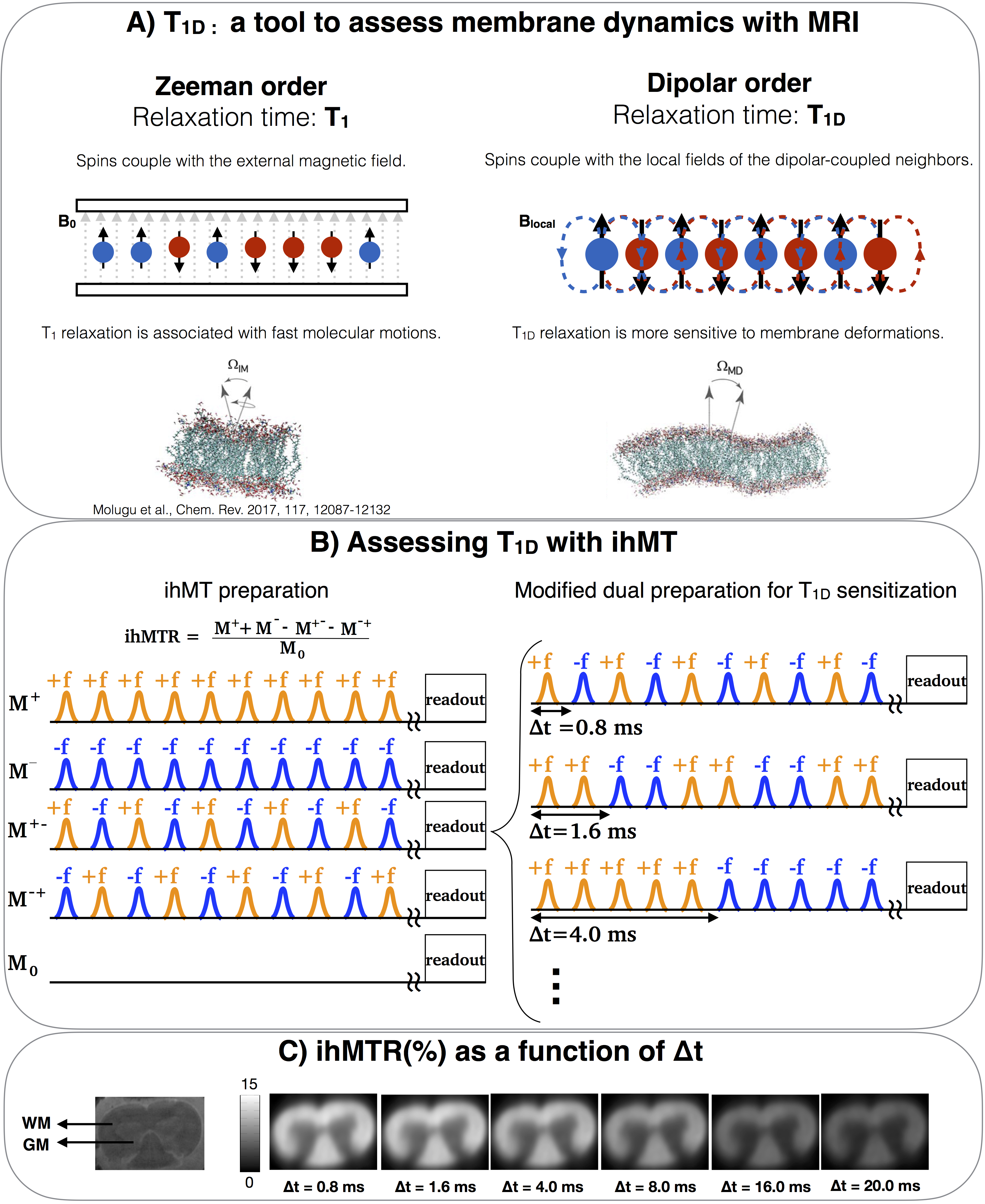
Figure 1.
A) Comparing T1 with T1D: T1 is the relaxation time of the Zeeman order and is associated with fast molecular motions in membranes. On the other hand, T1D is the relaxation time of the dipolar order and is also associated with slower and collective motions. Probing collective motions through T1D can be used to assess membrane integrity.
B)Modified ihMT preparation to assess T1D. The modification proposed in [9] allows the switching time, Δt, to be incremented, while keeping all other parameters constant.
C) ihMTR images of a rat spinal cord (ex-vivo) acquired with incremental Δt. The ihMTR attenuation is a function of T1D.
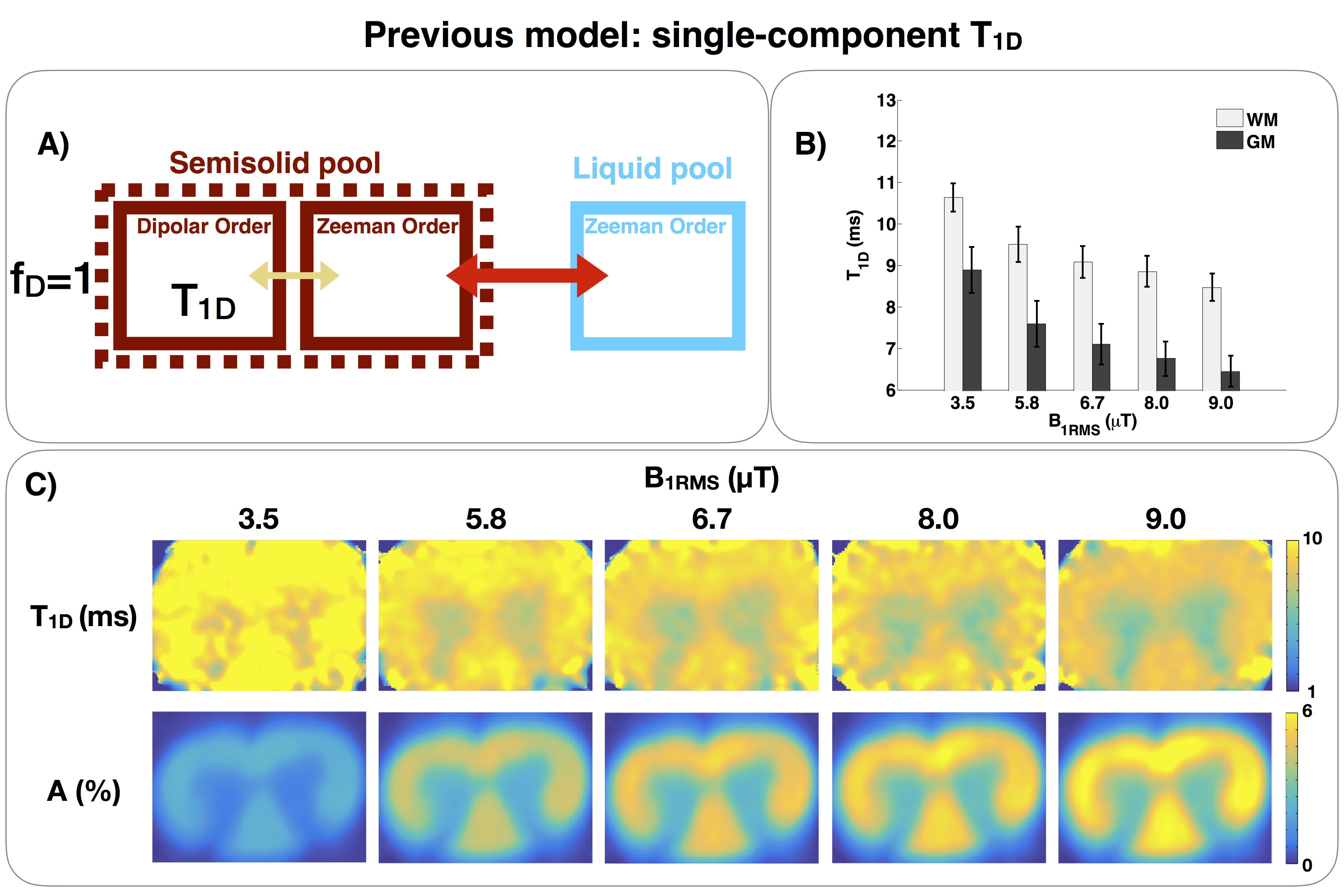
Figure 2.
A) Single-component T1D model of ihMT, with only one dipolar reservoir.
B) White matter (WM) and gray matter (GM) T1D fits of a rat spinal cord. Note that the apparent T1D decreases as B1RMS increases. This is likely explained because membranes exhibit multiple T1D components. As the irradiated RF power is increased, the short T1D components contribute more to ihMT. Since the single-component T1D model cannot account for this effect, it leads to a decrease of the apparent T1D.
C) T1D and A maps acquired with different B1RMS. A is the scaling factor9 of the function ihMTR(Δt).
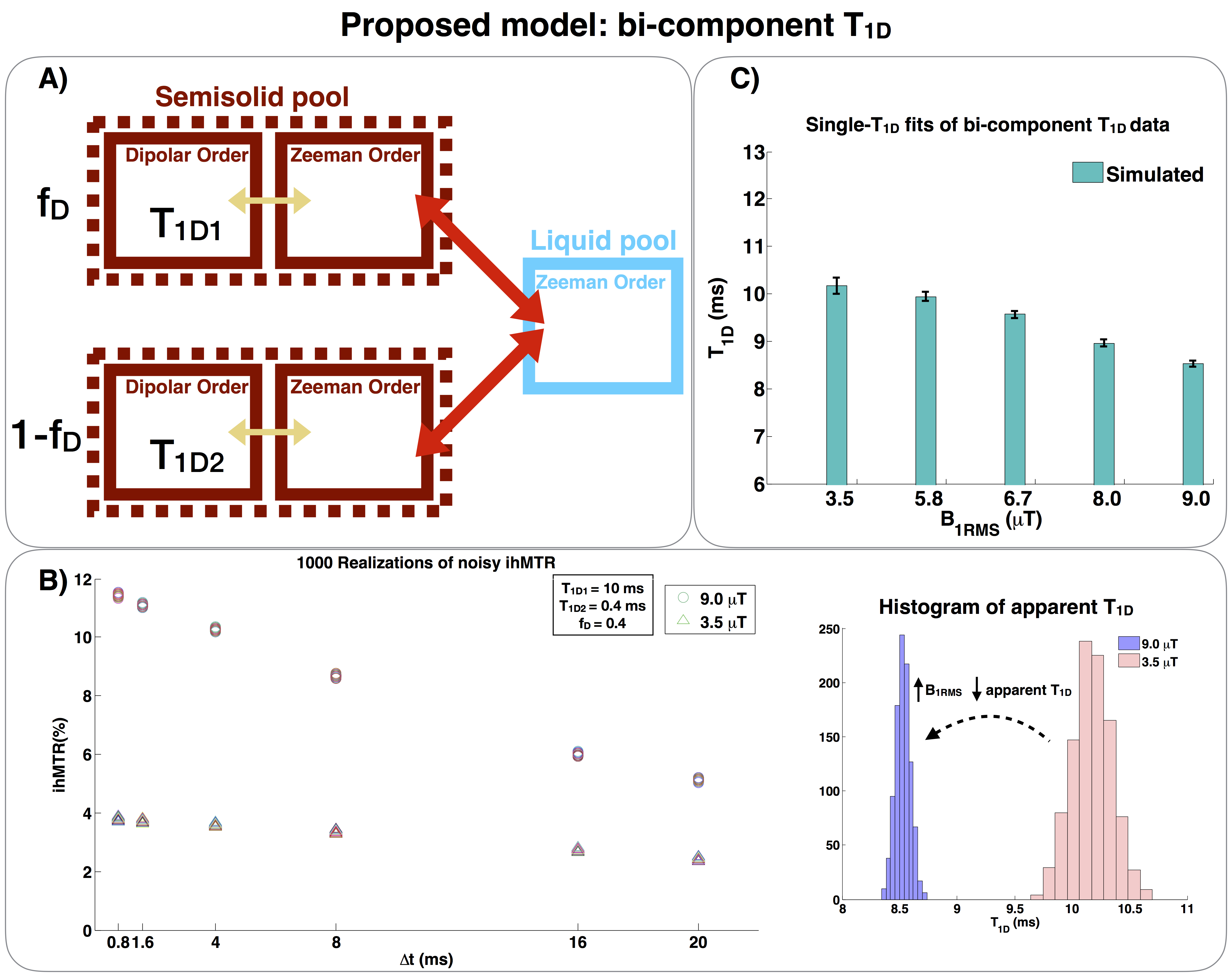
Figure 3.
A)Bi-component T1D model of ihMT, with two dipolar reservoirs. The fraction fD represents the fraction of the semisolid pool associated with T1D1, while 1-fD represents the fraction associated with T1D2.
B)Left: 1000 realizations of noisy ihMTR were generated with the proposed bi-component T1D model (T1D1=10 ms, T1D2=0.4 ms, fD= 0.4) for different B1RMSs. Right: Histograms of the apparent T1D distribution estimated by using the single T1D model (Fig. 2A) for B1RMS=3.5 and 9.0 μT.
C)Single-T1D fits (model of Fig. 2A) of bi-T1D simulated ihMTR data (model
of Fig.3A) for different B1RMSs. The apparent T1D decreases with B1RMS, as observed in the
experimental results.
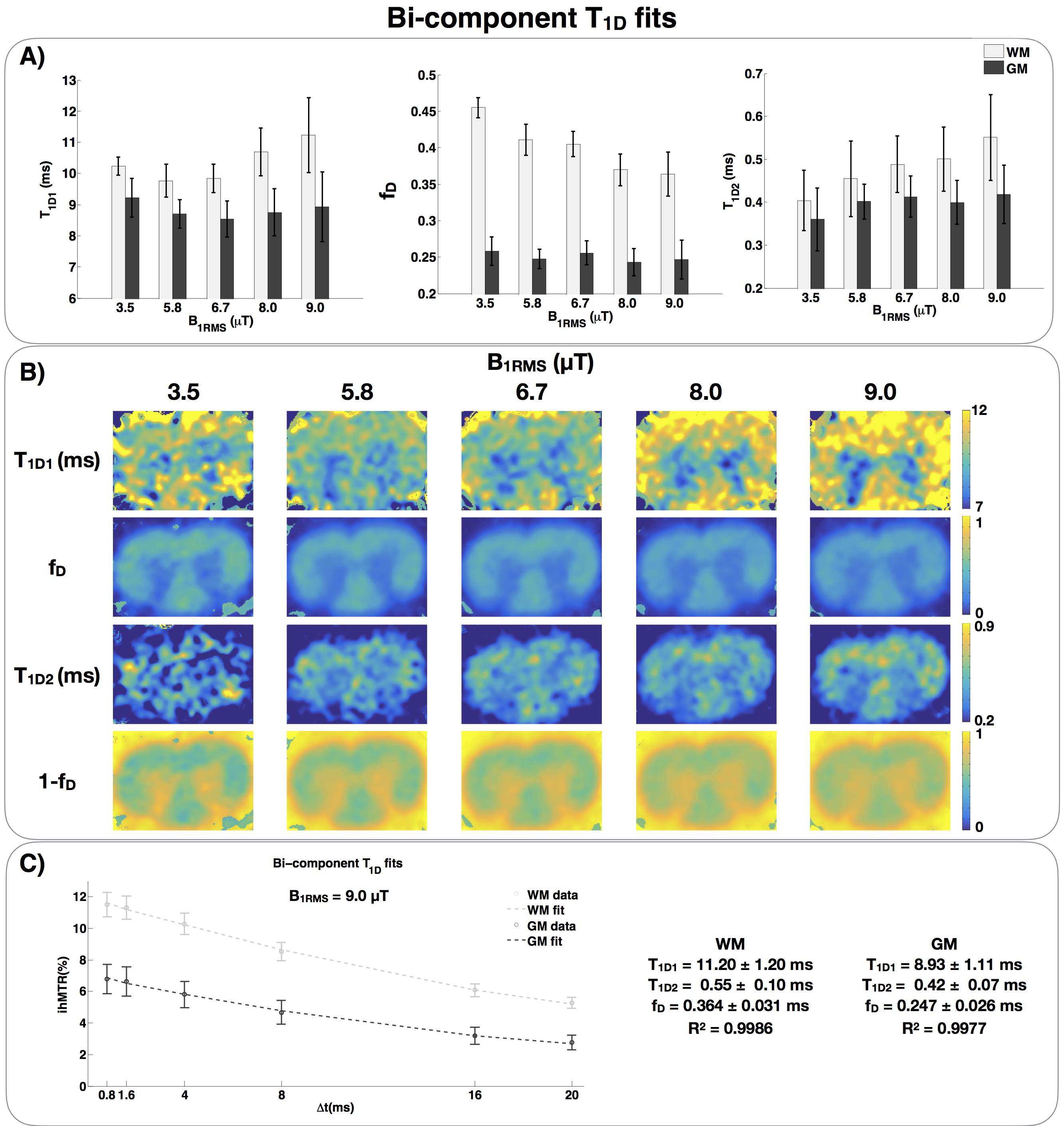
Figure 4.
A) Bi-component T1D fits of a rat spinal cord. T1D1 is in the order of 10 ms, and T1D2 is in the order of 400 μs. The fraction of long T1D, fD, is around 0.35-0.45 in WM and stable around 0.25 in GM. One can observe that the fraction of short T1D, 1-fD, increases in WM, suggesting that short T1D components are revealed.
B) T1D1, fraction fD, T1D2 and fraction 1-fD maps acquired with different B1RMS. Observe that the T1D2 map, the short T1D component, is revealed as B1RMS is increased.
C) Example of curve fit with the proposed bi-component T1D model.
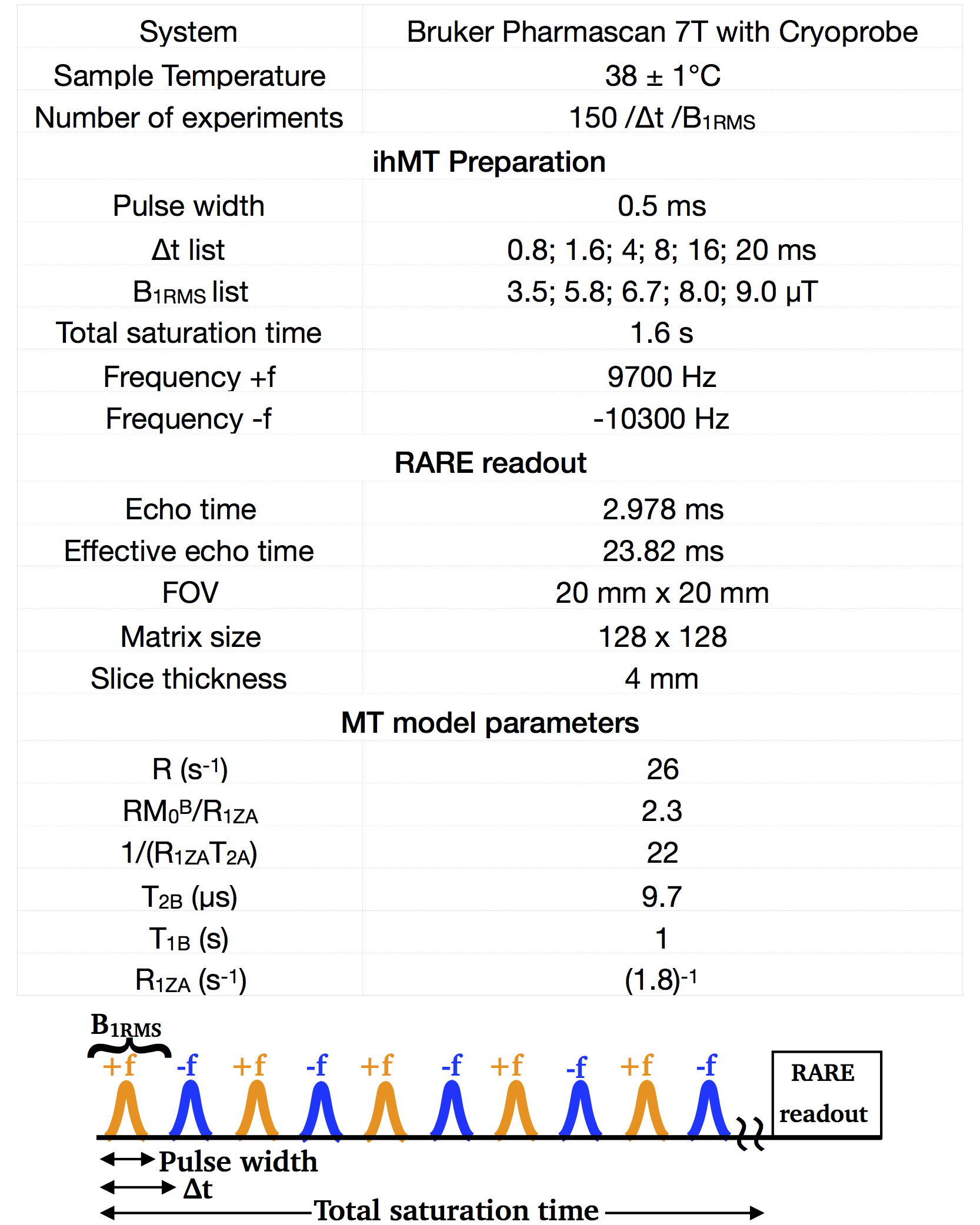
Table 1. Experimental parameters. The MT model parameters was based on [9].