4548
Robust 3D Bloch-Siegert based B1+ mapping using Multi-Echo General Linear Modelling1Wellcome Centre for Human Neuroimaging, UCL Institute of Neurology, London, United Kingdom, 2School of Biomedical Engineering & Imaging Sciences, King’s College London, London, United Kingdom
Synopsis
Robust quantification of the longitudinal relaxation rate (R1)—a widely used proxy marker of myelin content—requires highly accurate and precise estimation of the RF transmit field (B1+). The Bloch-Siegert shift (BSS) is a B1+-mapping method that allows calibration data to be acquired with the same spoiled gradient-echo readout used for variable flip angle R1 mapping. Here we show that systematic differences in steady state phase, caused by the interleaved nature typically adopted, lead to bias or loss of precision, but that these effects can be corrected for using a multi-echo approach and GLM fitting to isolate the BSS phase.
Introduction
The longitudinal relaxation rate (R1) is a useful marker of myelination enabling study of the structure-function relationship1,2. To achieve robust quantification and sensitivity to subtle inter-individual differences, highly accurate and precise estimation of the transmit field (B1+) is required. The Bloch-Siegert Shift (BSS)3 is a B1+-mapping procedure that uses off-resonance pulses to impart B1+-dependent phase. This phase can be isolated by acquiring two images using BSS pulses of opposite off-resonance frequency. The acquisition of these images is typically interleaved to increase robustness to long term phase inconsistencies, e.g. subject motion4 or scanner drift5. However, interleaving introduces an additional phase difference between the two acquisitions that causes bias in the B1+ map. These effects can be corrected by extracting the BSS-specific phase using a multi-echo acquisition with a general linear model (GLM) estimation procedure6
Methods
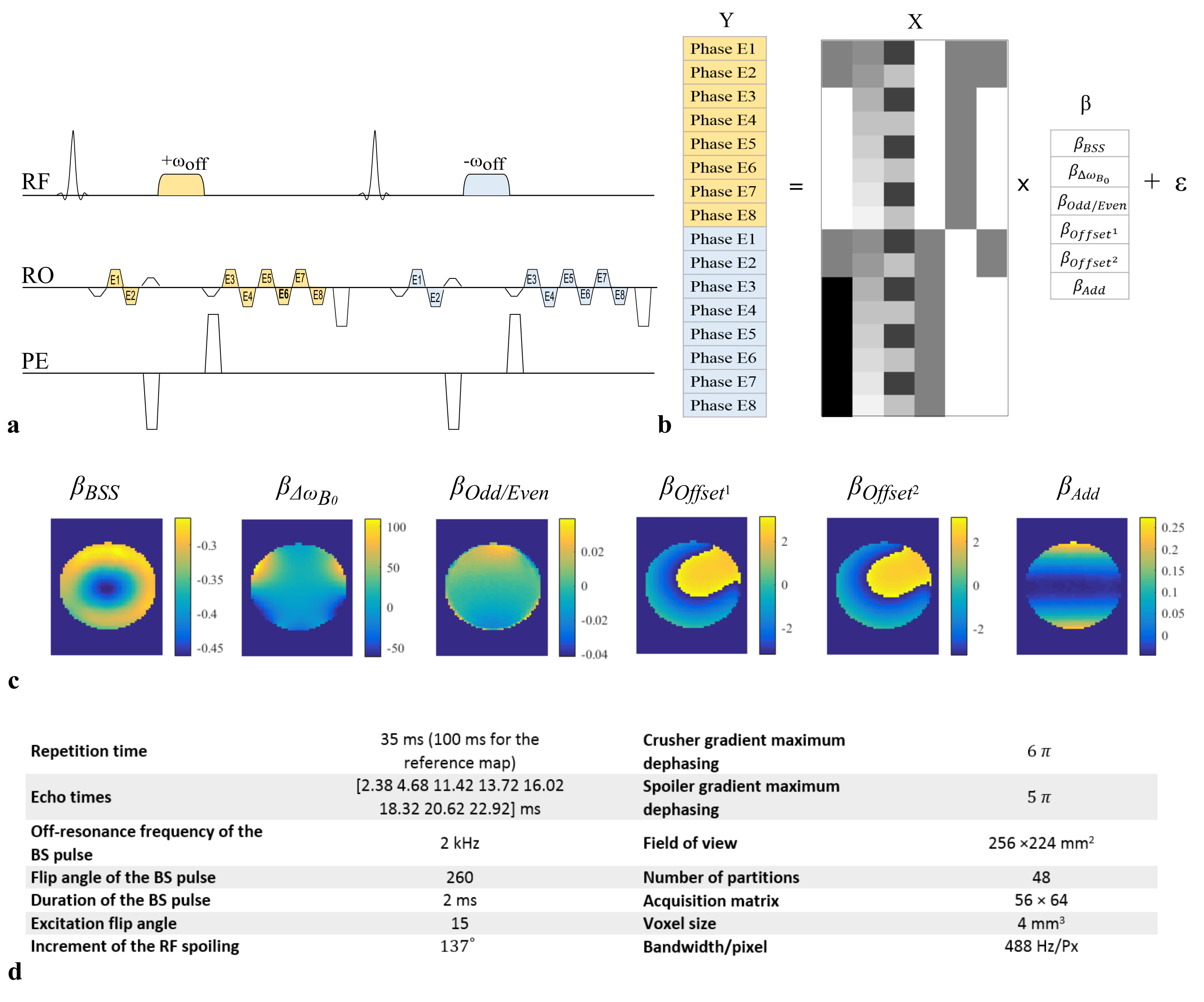
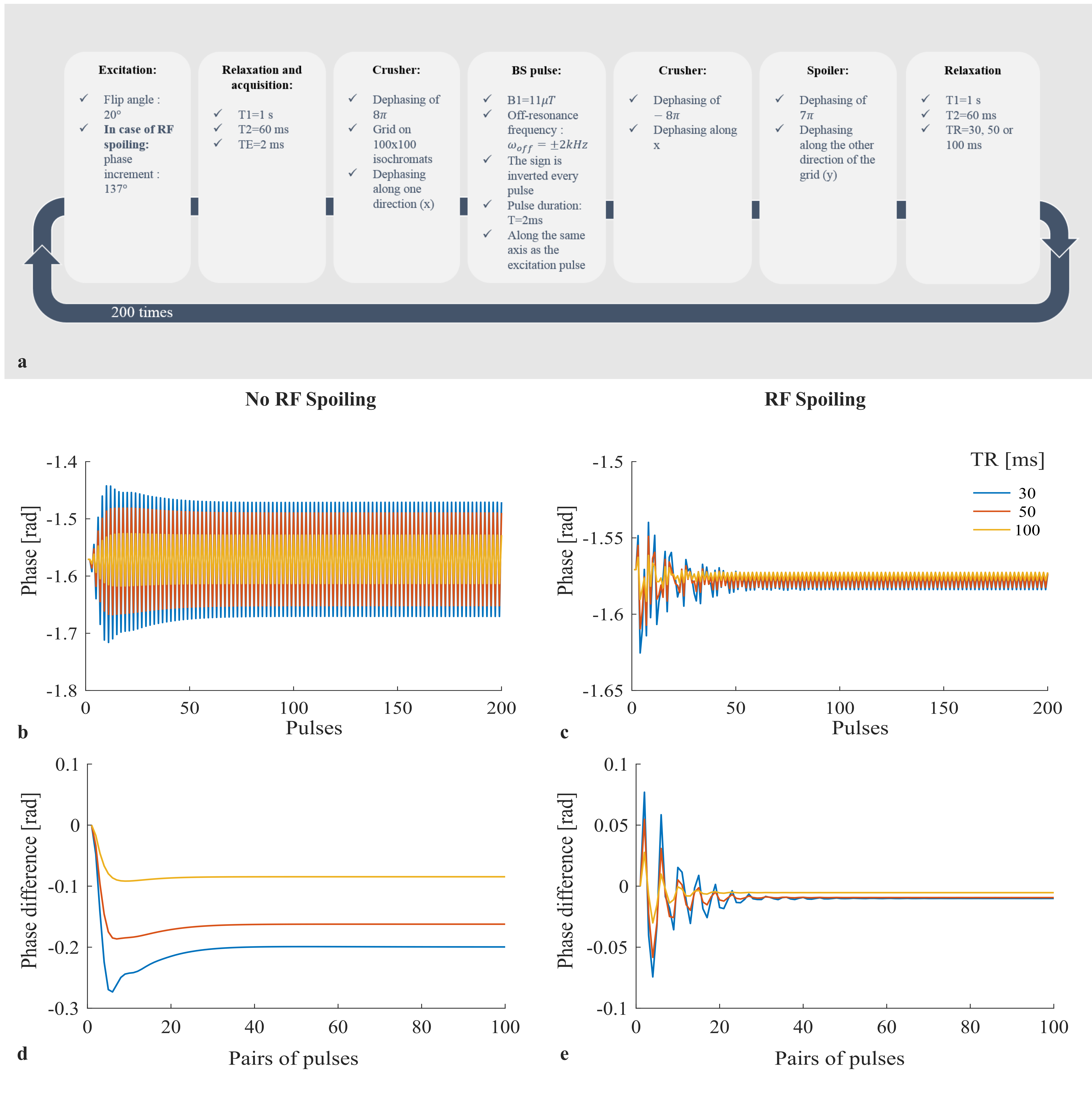
The phase of the steady-state signal of an interleaved BS acquisition (Fig.1a) was simulated as a function of pulse number using the procedure outlined in Fig.2a. The difference between opposite off-resonance frequency readouts prior to the BS pulse was also calculated. This was done to isolate any phase difference present from shot-to-shot that was not induced by the BSS. The impact of RF spoiling and TR was investigated. Phantom and in vivo experiments were conducted to verify the simulation results (protocol details in Fig.1d). All B1+ maps were reconstructed in real-time using Gadgetron7. These maps were calculated in two ways:
-The “classic” approach estimates B1+ efficiency from the phase difference between the echoes immediately following the BS pulses of opposite polarity (one measurement per BS-pulse frequency): $$$ \Phi_{BSS}=\frac{\Phi_{Diff}}{2}$$$
-The “GLM” approach isolates the phase specific to the BSS by fitting a GLM to the multi-echo data (Fig.1b,c): $$$\Phi_{BSS}=\beta_{BSS}$$$
$$$\Phi_{BSS}$$$ was then used to estimate B1+ ($$$B_1^+\propto \sqrt{\Phi_{BSS}}$$$).
For each estimation technique and protocol, errors were calculated with respect to a reference B1+ map acquired with TR=100ms and RF spoiling.
Results
Numerical simulations showed that the alternating off-resonance frequency of the BS pulse introduced an alternating phase across pulses(Fig.2b-c) that resulted in a phase discrepancy between the steady state of interleaved TRs (Fig.2d-e). This phase difference is present prior to the application of the BS pulse and will therefore manifest as a bias in the B1+ estimation when using the “classic” approach. This bias is predicted to be present with or without RF spoiling, but to be smaller using RF spoiling and/or with longer TR.
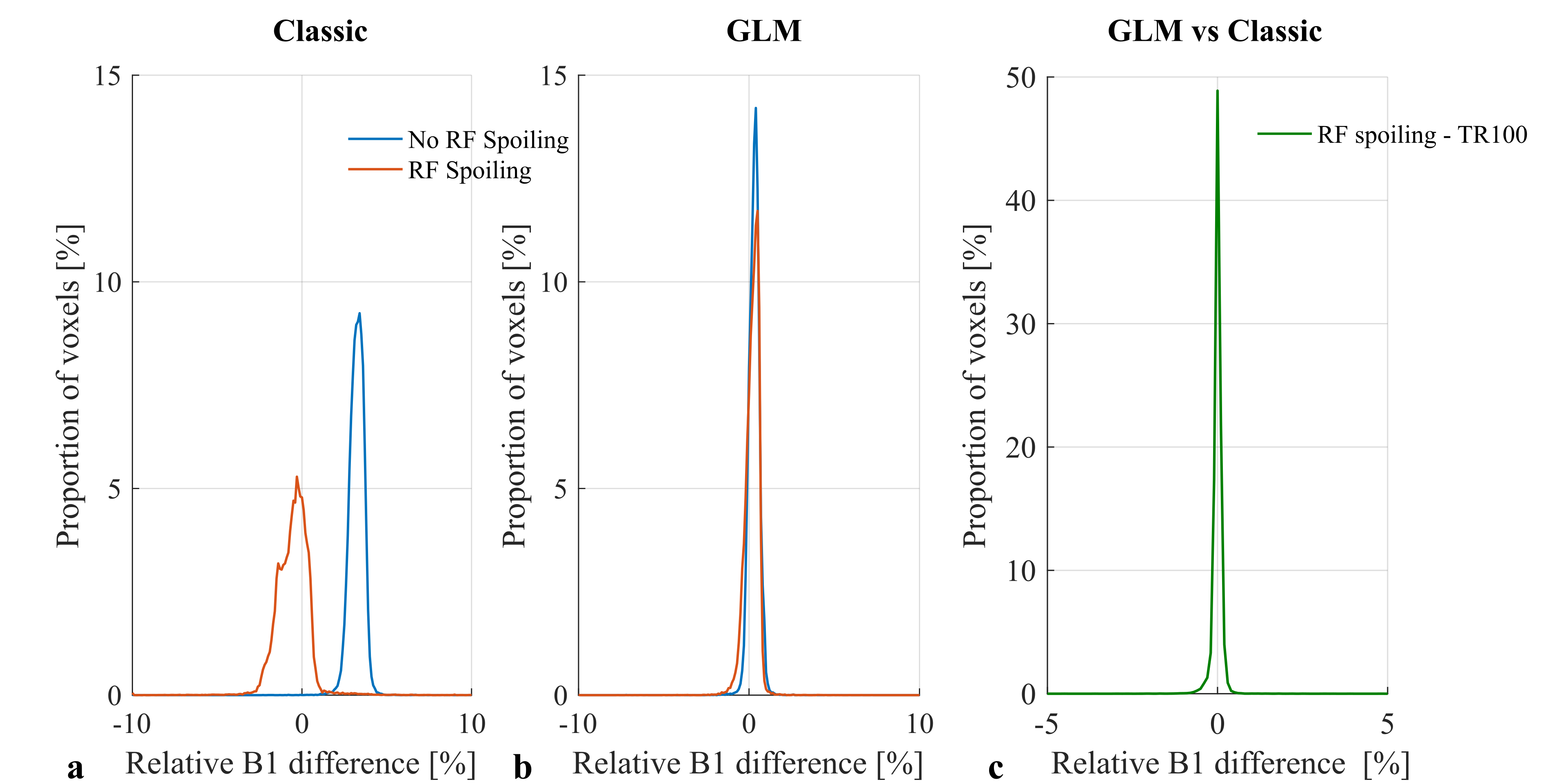
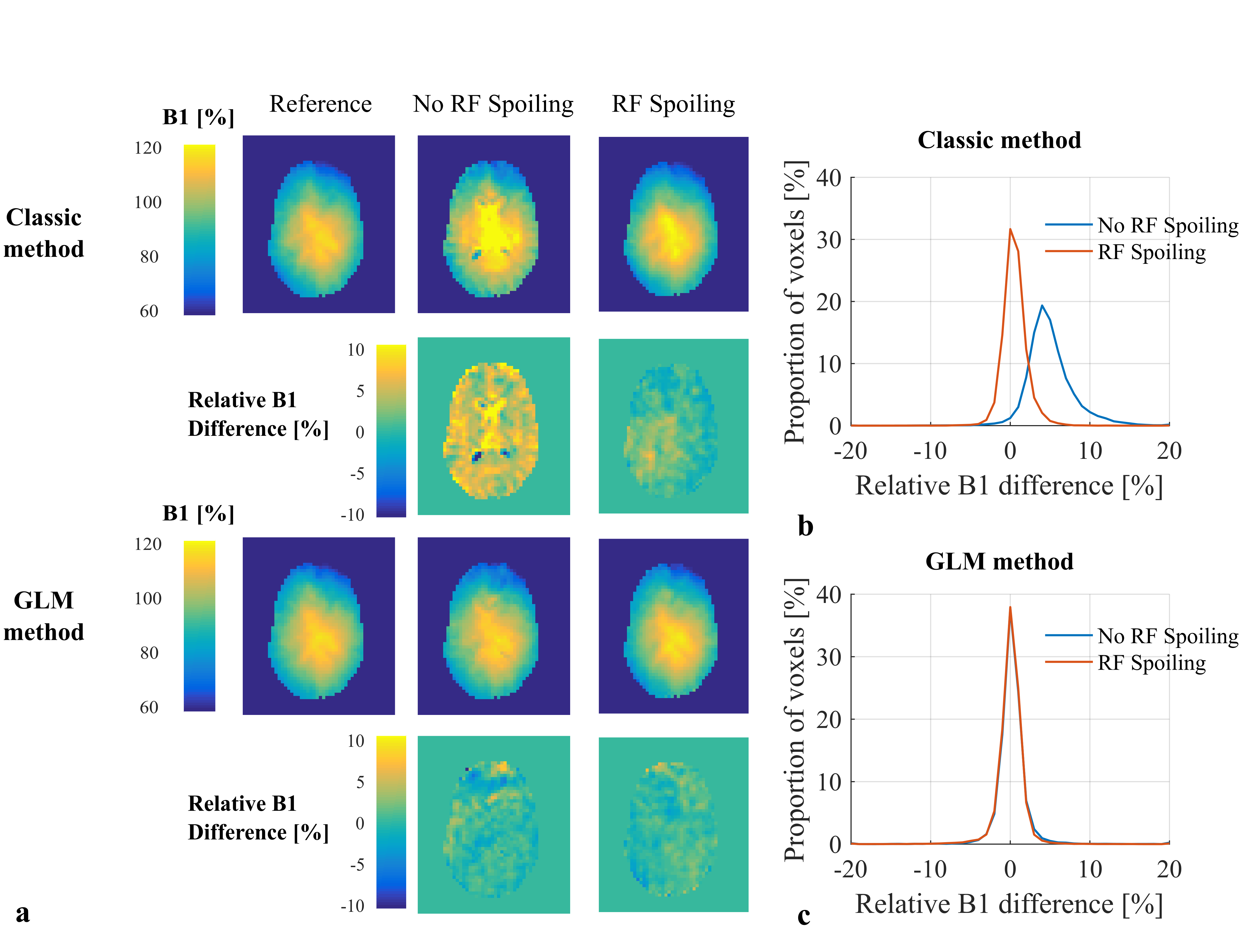
The results of phantom experiments were consistent with this. “Classic” B1+ maps contained bias when RF spoiling was not used (Fig.3a) that was greatly reduced by using RF spoiling. However, an additional source of variability, not predicted by the simulations, led to a broadening of the B1+ error histogram in the latter case. These effects (both bias and variance) were corrected by the GLM approach (Fig.3b). B1+ bias was also observed in vivo when not using RF spoiling (Fig.4a-b). In agreement with the phantom experiments, the GLM approach reduced both B1+ bias and variance with respect to the long-TR reference measurement.
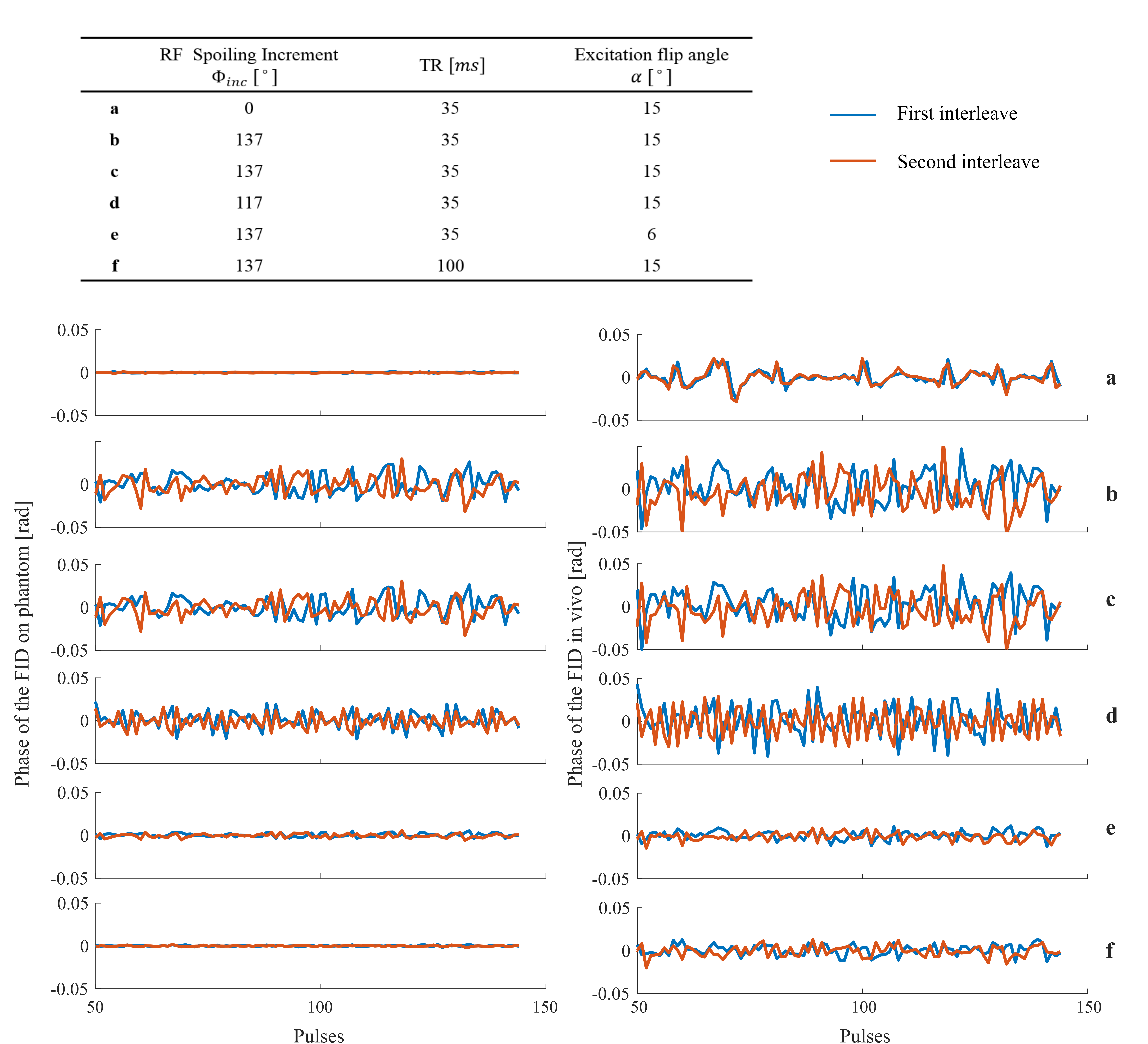
We hypothesised that the increased variance observed when using RF spoiling may be due to imperfect specification of the RF phase. To test this hypothesis, we acquired FID signals (with the same sequence, but only playing out the excitation pulse and spoiler gradient) in vivo and on phantoms using different RF spoiling increments (0°,117°,137°), TRs (35ms,100ms) and excitation flip angles (6°,15°). We computed the FID phases as a function of pulse number for the two interleaves. Consistent with our hypothesis, both in vivo and phantom experiments confirmed that phase variability observed with RF spoiling is deterministic, dependent on the RF spoiling increment, can be decreased with increasing TR or reducing the excitation flip angle and induces a spurious phase difference between the two interleaves.
Discussion
In this study we found that the classic BSS approach suffers from large B1+ biases (without RF spoiling) and increased variance (with RF spoiling). The GLM approach robustly isolated the B1+-specific phase leading to more precise and unbiased estimation.Conclusion
B1+ mapping with interleaved BS-pulses offers greater robustness to motion and scanner drift. However, the systematic difference of the BS-pulse frequency across TRs leads to a systematic difference in the steady-state phase for the two conditions that cannot be distinguished from the B1+-specific phase using the classic BSS approach. We demonstrated that this effect can be reduced using RF spoiling, but at the cost of sensitivity to any errors in pulse phase definition. Using a multi-echo acquisition and fitting a GLM offers robustness to both effects, enabling precise and accurate transmit field mapping, which is crucial for quantitative MRI.Acknowledgements
The Wellcome Centre for Human Neuroimaging is supported by core funding from the Wellcome [203147/Z/16/Z].References
1. Dick, F. K. et al. Extensive Tonotopic Mapping across Auditory Cortex Is Recapitulated by Spectrally Directed Attention and Systematically Related to Cortical Myeloarchitecture. J. Neurosci. 37, 12187–12201 (2017).
2. Carey, D., Krishnan, S., Callaghan, M. F., Sereno, M. I. & Dick, F. Functional and Quantitative MRI Mapping of Somatomotor Representations of Human Supralaryngeal Vocal Tract. Cereb. Cortex N. Y. NY 27, 265–278 (2017).
3. Sacolick, L. I., Wiesinger, F., Hancu, I. & Vogel, M. W. B1 Mapping by Bloch-Siegert Shift. Magn. Reson. Med. Off. J. Soc. Magn. Reson. Med. Soc. Magn. Reson. Med. 63, 1315–1322 (2010).
4. Kameda, H. et al. Improvement of the repeatability of parallel transmission at 7T using interleaved acquisition in the calibration scan. J. Magn. Reson. Imaging 48, 94–101 (2018).
5. Lesch, A., Petrovic, A. & Stollberger, R. Robust implementation of 3D Bloch Siegert B1 mapping. in (2015).
6. Corbin, N., Acosta-Cabronero, J., Weiskopf, N. & Callaghan, M. F. Rapid B1 mapping based on the Bloch-Siegert shift using a single offset frequency and multi-echo readout. in (2018).
7. Hansen, M. S. & Sørensen, T. S. Gadgetron: An open source framework for medical image reconstruction. Magn. Reson. Med. 69, 1768–1776 (2013).
8. Duan, Q., van Gelderen, P. & Duyn, J. Improved Bloch-Siegert Based B1 Mapping by Reducing Off-Resonance Shift. NMR Biomed. 26, 1070–1078 (2013).
Figures