4461
The in vivo impact of diffusion spoiling on the estimate of T1 using spoiled gradient echoes with variable flip angles1Wellcome Centre for Human Neuroimaging, UCL Institute of Neurology, London, United Kingdom, 2School of Biomedical Engineering & Imaging Sciences, King’s College London, London, United Kingdom
Synopsis
Incomplete spoiling of the transverse MRI signal causes errors in the T1 time estimated from variable flip angle measurements acquired with spoiled gradient-echo images. Diffusion spoiling is thought to lessen these effects. However, these conclusions are based on phantom experiments, using very long T2 times, or from in vivo simulation using infeasibly strong diffusion spoiling. Here we perform simulation and in vivo experiments to characterise the impact of diffusion spoiling in the short T2, low spoiling regime. We show that even under these conditions, diffusion spoiling reduces the dependence of the estimated T1 on both the phase-increment and the transmit field.
Introduction
Combining spoiled gradient echo datasets acquired with different flip angles enables high resolution mapping of the longitudinal relaxation time (T1), achieving whole brain coverage in feasible scan durations. However, this approach relies on complete spoiling of the transverse magnetisation between successive excitations. Otherwise, the apparent T1 estimated from the Ernst equation will not match the true value. This effect has been characterised and a correction scheme proposed1. Diffusion effects2 have also been shown to affect the estimated T1. However, this effect has only been explored using simulation and experiment with very long T2 but moderate spoiler moment, or, in simulation only, using in vivo T2 times but with infeasibly large spoiler moments (250-1000mT.ms/m). Here we characterise the interaction between the choice of RF spoiling increment and the amplitude of the spoiler moment via simulation and in vivo experiment using feasible spoiler moments.Methods
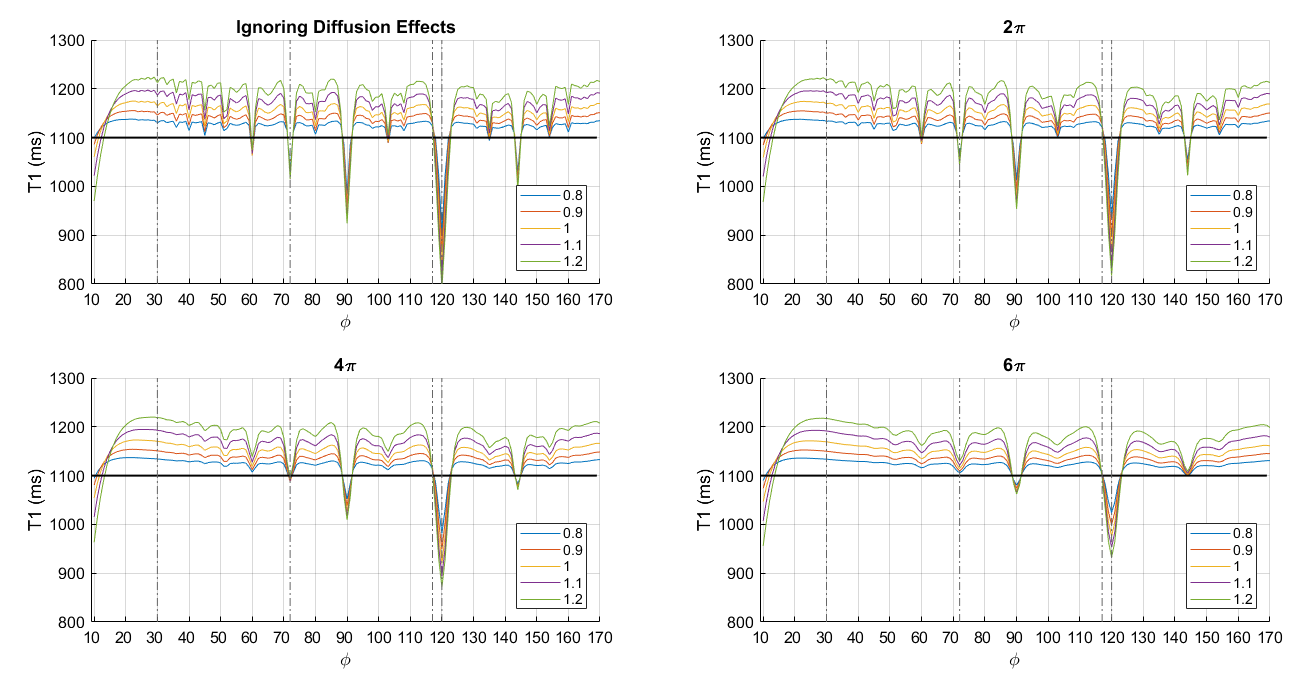
Simulations were performed using the EPG formalism (http://mriphysics.github.io/), which can optionally include diffusion effects. The simulations used T1=1100ms, T2=100ms (estimated in a separate in vivo T2 mapping experiment), and a diffusion coefficient2 of 0.8x10-9m2s-1. The simulations tested a range of B1+ efficiencies (80-120%) and RF spoiling increments (10-170°). The simulations were performed (a) without diffusion effects, and (b) accounting for the full multi-echo readout (8 echoes) and spoiler gradients (applied on the same axis) that were used for the in vivo experiments. Total dephasing moments per TR of 2, 4 or 6π across one voxel were simulated. The spoiler gradient had a constant amplitude of 33.3mT/m and its duration was increased to achieve the different spoiling.
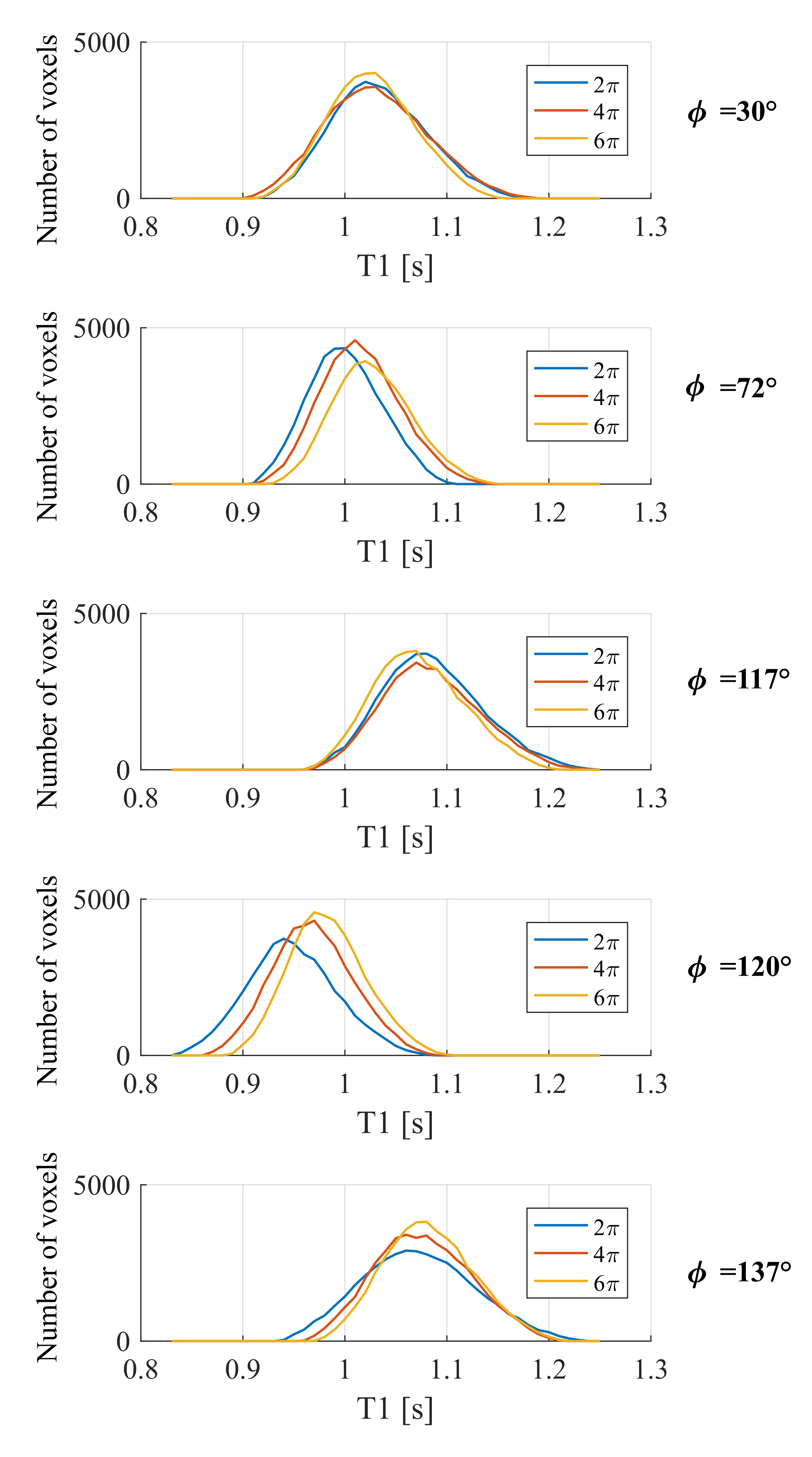
In vivo data were acquired on a Siemens Prisma 3T scanner using a 32-channel head coil for signal reception and the body coil for signal transmission. Three spoiling conditions (net dephasing per TR of 2, 4 or 6π across a voxel), and five RF spoiling increments (30°,72°,117°,120° and 137°) were tested. Additional calibration data to map the B1+ efficiency were acquired using an established spin-echo/stimulated-echo acquisition3. Two FLASH volumes were acquired with flip angles of 6° and 21° (TR=22ms) with a slab-selective excitation and field-of-view of 240x180x56mm3 with 1mm isotropic resolution. The TE ranged from 2.2ms to 18.3ms in 2.3ms steps. The hMRI toolbox (hMRI.info) was used to estimate T1 from these data. No correction for imperfect spoiling was applied. Histograms of T1 values in white matter (p(WM)>0.9) from the central 7 slices of the slab were computed.
Results
Simulations predicted a strong dependence of the T1 estimate on the moment of the spoiler gradient, even for these moments that can feasibly be achieved in vivo. The simulation ignoring diffusion effects is highly featured (Fig.1). The T1 is generally over-estimated and there is considerable variability with distinct troughs where the T1 estimate falls dramatically, e.g. at 72, 90, 120 and 135°. The nuances of these curves are supressed by incorporating diffusion effects, with greater suppression as the spoiler moment is increased.
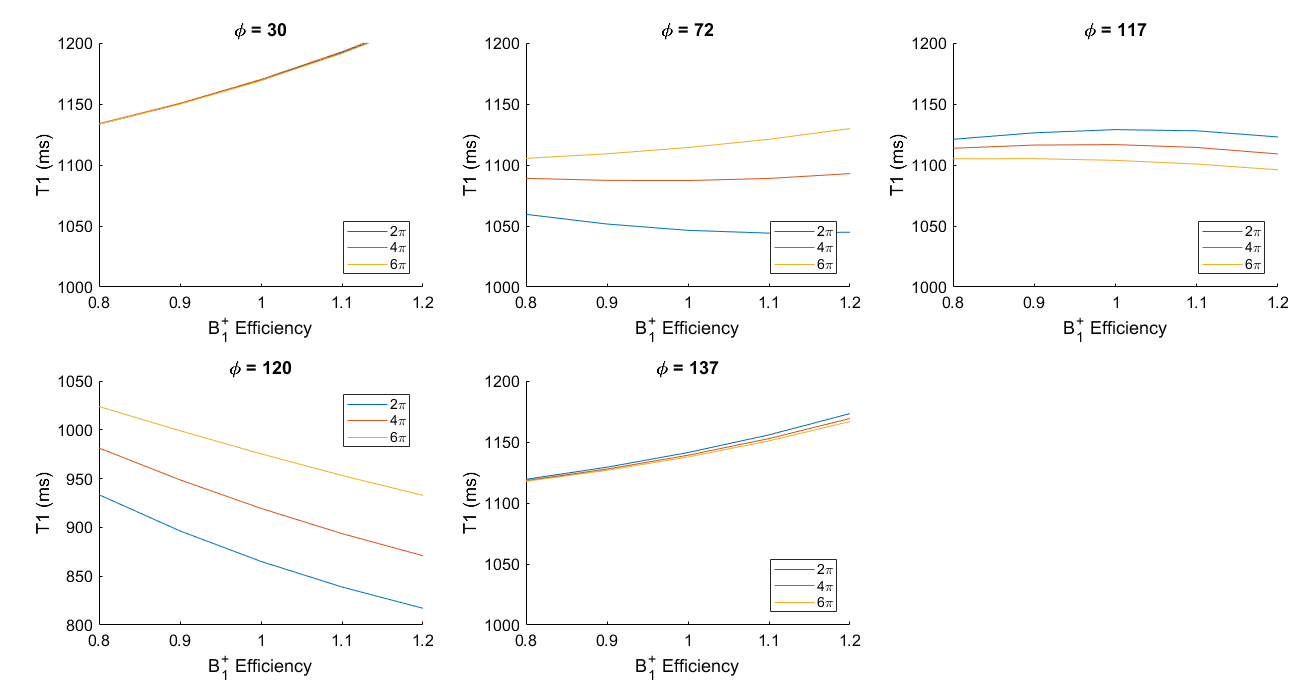
The degree of diffusion-induced spoiling also impacts the B1+ dependence of the T1 estimate, which is predicted to vary considerably with RF spoiling increment (Fig.2).
In accordance with the simulations, the mean T1 value over the white matter was strongly dependent on the RF spoiling increment used (Fig.3). The mean T1 estimates obtained with increments of 72° and 120° were biased to lower values than 30°, 117° or 137°. Consistent with the simulations, this bias was decreased as the spoiler moment increased, while no substantial change in the average T1 values was observed for increments of 30°, 117° or 137°.
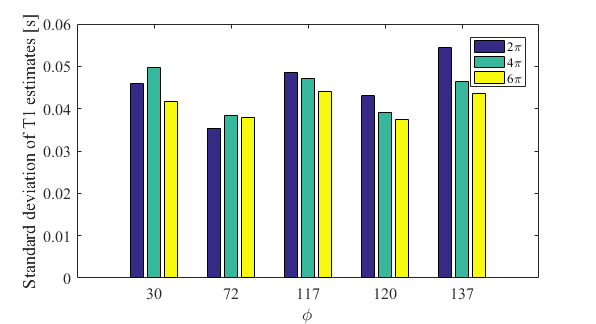
The T1 estimates are also predicted to have a strong dependence on the B1+ efficiency. Since the anatomical variability is consistent across all conditions, we interpret the width of the histogram as capturing the degree of B1+ dependence, with broader histograms indicating greater B1+ dependence. As the spoiler moment increased, the standard deviation of the T1 estimates decreased for 117°, 120° and 137° (Fig.4). For increments of 30° and 72° this was only true when going from 4 to 6π.
Discussion
Even with spoiler moments that can feasibly be achieved in in vivo imaging, the imparted diffusion spoiling has a considerable impact on the estimated T1 and interacts strongly with the chosen RF spoiling increment.
Of note, these results are specific to the multi-echo readout used here, which will impart more diffusion spoiling than previously-examined single-echo scenarios. Furthermore, the specific form of the simulations will depend on the parameters used (e.g. T1, T2 and diffusion coefficient). These choices may underlie the observed differences with respect to the in vivo experiments.
Conclusions
Our findings indicate that the impact of diffusion spoiling should be considered when optimising T1 mapping protocols using spoiled gradient echo acquisitions.Acknowledgements
The Wellcome Centre for Human Neuroimaging is supported by core funding from the Wellcome [203147/Z/16/Z].
References
1. Preibisch, C. & Deichmann, R. Influence of RF spoiling on the stability and accuracy of T1 mapping based on spoiled FLASH with varying flip angles. Magn. Reson. Med. 61, 125–35 (2009).
2. Yarnykh, V. L. Optimal radiofrequency and gradient spoiling for improved accuracy of T1 and B1 measurements using fast steady-state techniques. Magn. Reson. Med. 63, 1610–26 (2010).
3. Lutti, A. et al. Robust and Fast Whole Brain Mapping of the RF Transmit Field B1 at 7T. PLoS One 7, e32379 (2012).
Figures