4404
Myelin Water Imaging Profiles Along White Matter Tracts1School of Biomedical Engineering, The University of British Columbia, Vancouver, BC, Canada, 2Department of Medicine, Division of Neurology, The University of British Columbia, Vancouver, BC, Canada, 3Department of Radiology, The University of British Columbia, Vancouver, BC, Canada, 4Department of Physics & Astronomy, The University of British Columbia, Vancouver, BC, Canada
Synopsis
Myelin water fraction (MWF) maps are spatially noisy. Here we investigated a possible inherent spatial structure of MWF values along diffusion tensor imaging (DTI)-derived white matter (WM) tracts in 41 healthy subjects. Sixteen major fibre bundles were extracted and MWF was computed in sub-segments along each fibre tract and compared to surrounding voxels. MWF values were more spatially coherent along fibre bundles than elsewhere. The profile along the trajectory of fibre bundles estimated subjects’ age more accurately than tract-averaged MWF. We conclude that the spatial MWF distribution in WM consistently follows a distinct pattern along underlying fibre bundles across subjects.
Introduction
Much research focuses on assessing myelin within specific local regions or entire fibre bundles. However, imaging-based myelin features, e.g. myelin water fraction (MWF), may be influenced by a number of factors such as disease processes 1, changes along white matter (WM) pathways due to myelogenesis 2, or changes due to aging.
In this work we aim to characterise the basic spatial structure of MWF maps with respect to WM microstructural organisation. We hypothesize that the MWF variance along major WM fibre bundles will be lower than in other regions of the brain. Furthermore, we hypothesize that different major WM tracts have a characteristic spatial pattern of MWF values allowing for identification of local changes that may be of diagnostic value.
Methods
MRI data was collected for 41 healthy subjects (23F), median age 28yrs (range 20-52). All data were acquired on a Philips Achieva 3T MRI with an 8 channel head coil. T2 relaxation data were collected using a modified gradient and spin echo (GRASE) sequence with 32 echoes, TE=10ms, TR=1000ms, voxel size 1x1x2.5mm. Twenty-one subjects had diffusion tensor imaging (DTI) data sets acquired with 16 directions, b-value of 900 s/mm2 and one b0 volume. The remaining 20 subjects had DTI data sets with 32 directions, b-value of 700 s/mm2 and one b0 volume. The voxel size was 0.8x0.8x2.2mm for both sequences.
The multi-echo GRASE sequence was analysed as described in Prasloski, et al 3. DTI data were corrected for geometric distortions and subject motions using DTIFIT 4 prior to fitting the tensors and performing whole brain tractography. Sixteen major white matter fascicles were segmented with the AFQ toolbox 5. The first echo of the GRASE data was registered to each subject’s b0 image and the resulting transformation matrix was then applied to each MWF map using FLIRT 4.
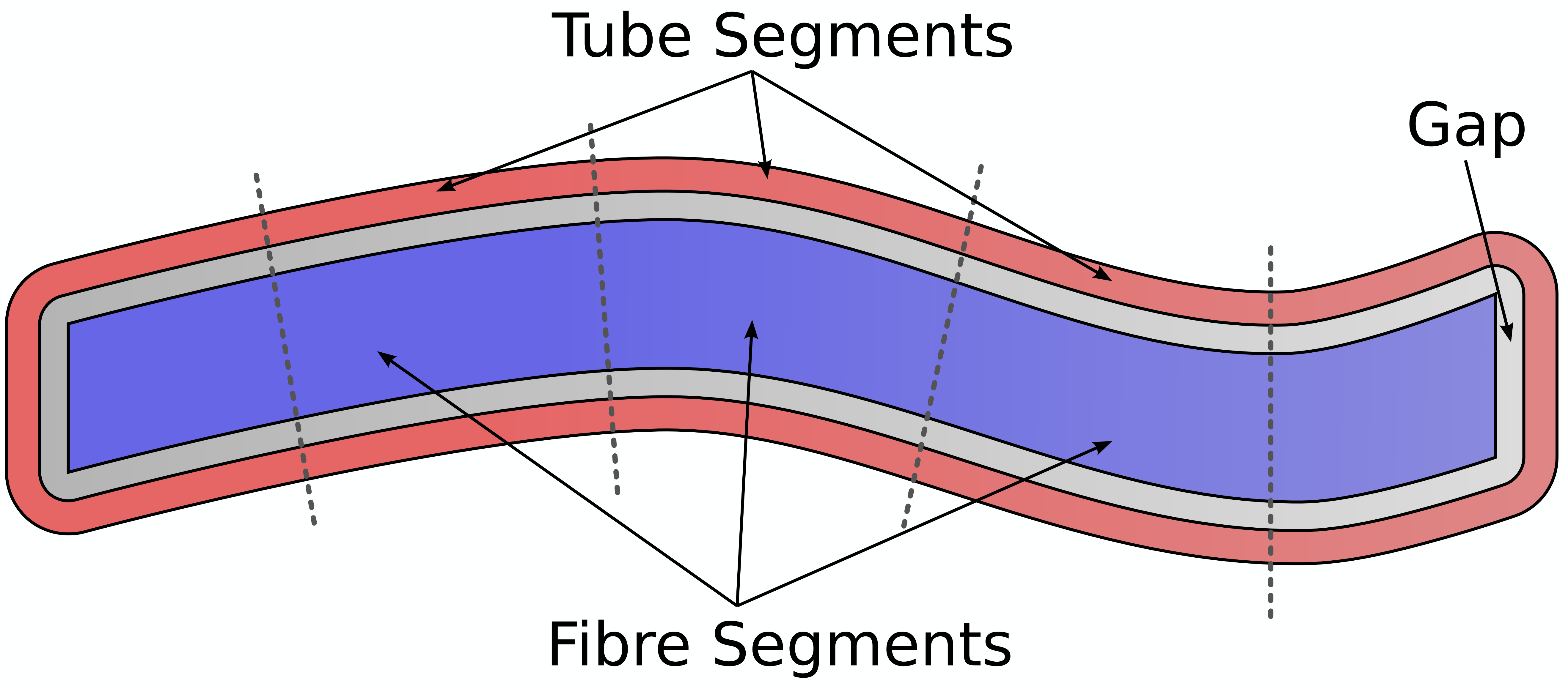
We compared the coefficient of variation (CoV) of MWF values in fascicles to their respective “tubes” – i.e. two voxel layer of WM voxels surrounding each fascicle (Figure 1).
Further, we compared the CoV gradient in 15 sub-segments along the fascicles to the CoV gradient in tube segments perpendicular to the fibre orientation.
Lastly, we performed linear regression to estimate subjects’ age using either the total average MWF of a fibre bundle or the average MWFs in sub-segments to determine the significance of MWF values along fibre bundles.
Reults
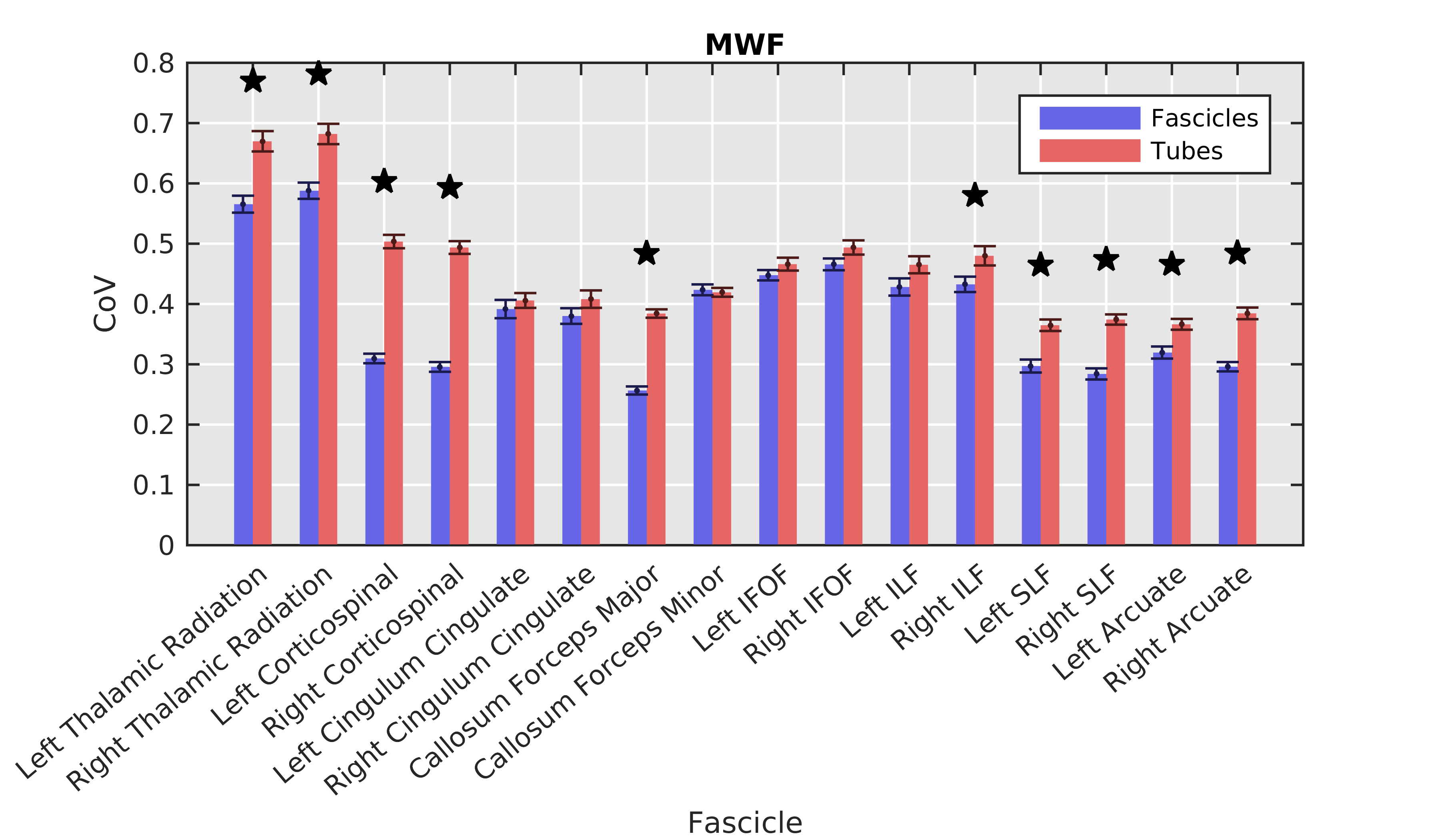
Fibre bundles demonstrated significantly (p<0.001) lower CoV than their respective tubes in the left/right corticospinal tract callosum forceps major, right inferior longitudinal fasciculus, left/right superior longitudinal fasciculus, and left/right arcuate (Figure 2).
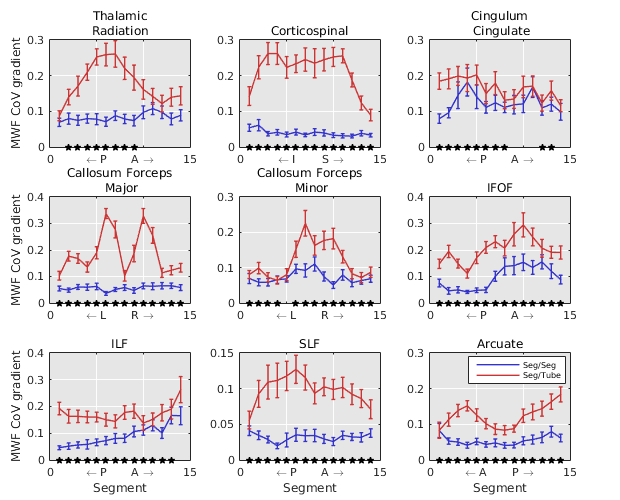
Comparing the CoV gradients along WM fibre bundles to a direction perpendicular revealed lower CoV gradients along fibre segments than perpendicular to segments (Figure 3).
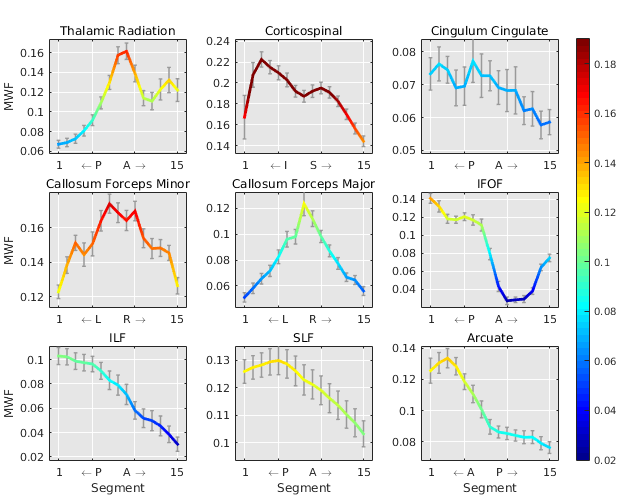
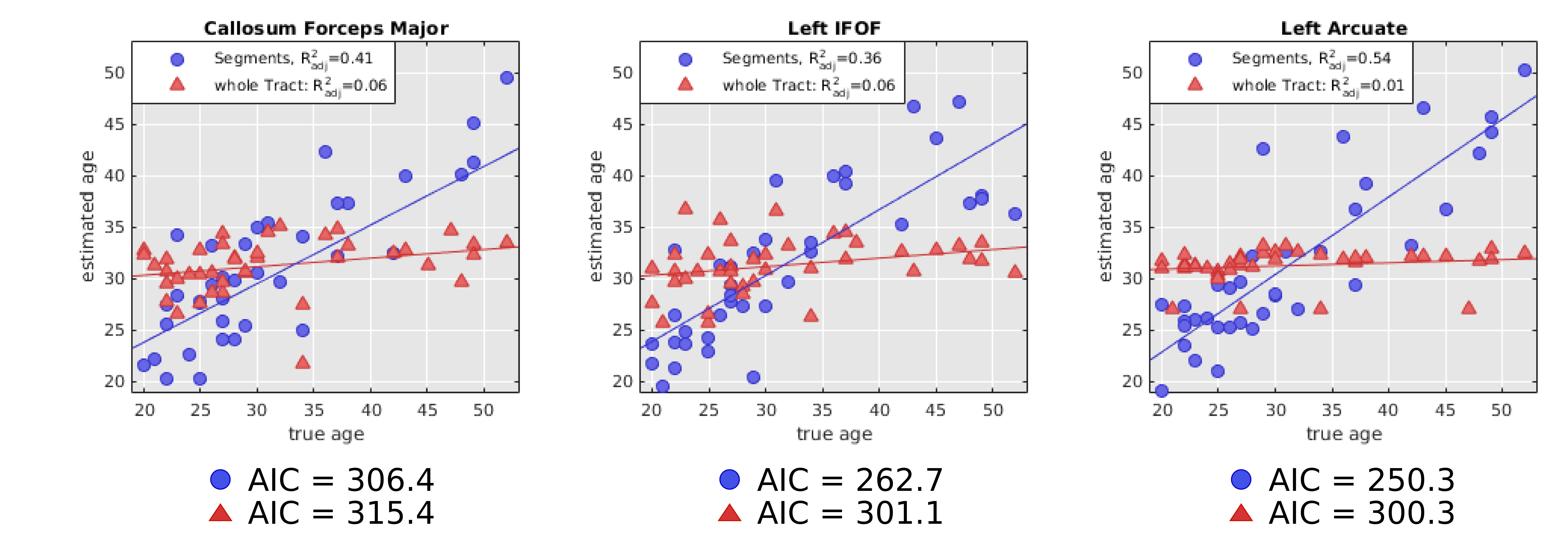
The average sub-segment MWFs along major fibre bundles are displayed in Figure 4 which shows a characteristic spatial pattern for each fibre bundle. A linear regression model for each fascicle, found the callosum forceps major (p<0.05), left IFOF (p<0.05), and left arcuate (p<0.01) to be significantly associated with age (Figure 5). The age estimation based on MWFs in fibre segments outperformed the estimation obtained using only the simple tract-average MWF per fascicle, as can be seen by lower AIC values in Figure 5.
Discussion
MWF values were more consistent along a fibre bundle compared to “tubes” surrounding the fascicles. Furthermore, MWF variance exhibited a spatial gradient in major WM fibre bundles which was lower along fibre tracts compared to perpendicular directions.
It is evident that each major fibre bundle has its own specific pattern of myelination, and a general trend from higher to lower MWF values from posterior to anterior can be seen. Some structures such as the callosum forceps major/minor have intricate patterns that reflect the brains subtle myelination texture. Those spatial patterns provide a more detailed view on WM tissue composition which could be exploited to pinpoint areas of myelin alterations, rather than averaging over whole fibre bundles which may mask subtle, local differences in myelination. This may be of use when one wishes to determine age effects on myelination or infer disease-related changes.
Conclusion
In conclusion, our results suggest the existence of a spatial pattern in MWF despite the noisy appearance of MWF maps. Likewise, when performing an ROI based analysis one should be aware of underlying white matter microstructure when investigating large ROIs that include different fibre tracts. When investigating whole fibre tracts, it may be inadvisable to integrate the MWF over the entire tract length rather than looking at smaller segments within each tract.Acknowledgements
We thank the MRI technicians and all participating subjects.References
1. Vargas W, Monohan E, Pandya S, et al. Longitudinal Changes in Myelin Water Fraction in New Multiple Sclerosis Lesions (S44.005). Neurology 2014;82.
2. Yakovlev P, Lecours A. The Myelogenetic Cycles of Regional Maturation of the Brain. Regional Development of the Brain in early Life. 1967.
3. Prasloski T, Mädler B, Xiang QS, et al. Applications of stimulated echo correction to multicomponent T2 analysis. Magn. Reson. Med. 2012;67(6):1803–1814.
4. Jenkinson M, Beckmann CF, Behrens TEJ, et al. FSL. Neuroimage 2012;62(2):782–90.
5. Yeatman JD, Dougherty RF, Myall NJ, et al. Tract Profiles of White Matter Properties: Automating Fiber-Tract Quantification. PLoS One 2012;7(11).
Figures