4323
Metabolism of hyperpolarized [1-13C]pyruvate in isolated perfused kidneysNesmine R Maptue1, Gaurav Sharma1, Miriam Trigo1, Alexander M Funk1, Thomas Hever1, Xiaodong Wen1, Craig R Malloy1, A. Dean Sherry1, and Chalermchai Khemtong1
1Advanced Imaging Research Center, University of Texas Southwestern Medical Center, Dallas, TX, United States
Synopsis
Kidney primarily oxidizes fatty acids and ketone bodies for ATP production but the substrate preference is altered in metabolic diseases. Renal metabolism is generally evaluated in kidney slices and in vivo. However, real-time metabolism has not been studied in isolated perfused kidneys where substrate availability and pharmacological interventions can be well-controlled. Here, we investigate pyruvate metabolism in isolated perfused rat kidneys by hyperpolarized 13C NMR. Results show that hyperpolarized 13C-pyruvate was rapidly metabolized in the kidneys and productions of 13C-bicarbonate, 13C-alanine, and 13C-lactate were detected. the utilization of pyruvate for energy production was confirmed by isotopomer analyses of tissue extracts.
Purpose
Kidneys are one of the most metabolically active and energy-demanding organs in the body. Fatty acids and ketone bodies are primary sources for renal energy production but other substrates can also be oxidized. Metabolic heterogeneity is present in the kidney with the cortex and medulla displaying metabolic differences. Noninvasive and sensitive imaging modalities that can accurately detected altered metabolism in the kidneys will certainly be very useful for the detection and treatments of metabolic diseases such as diabetes. In this study, we investigated substrate metabolism in isolated perfused rat kidneys by hyperpolarized 13C NMR. The goal of this study was to evaluate whether hyperpolarized 13C NMR can be used as a tool to interrogate real-time metabolism of 13C-enriched substrates in functioning perfused rats kidneys.Methods
[1-13C]Pyruvate doped with 15 mM trityl radical was polarized in a HyperSense. Sprague-Dawley rats (500-600 g) were used for this study. Abdominal midline incision was made under a general anesthesia (1.5% isoflurane in oxygen). The abdominal aorta was cannulated and the right kidney was isolated. The kidney was then connected to the perfusion apparatus (Fig. 1A) and perfused with a non-circulating modified Krebs-Henseleit buffer (KH, pH=7.4) containing glycine (2.3 mM) and cysteine (0.5 mM) to maintain the vitality of the kidney by increasing the glomerular filtration rate during the perfusion. Oxidizable substrates available to the kidney in the perfusate were 2mM [U-13C]pyruvate (group 1) or 2mM [U-13C]pyruvate and 0.2 mM [2,4,6,8-13C]octanoate (group 2). Four kidneys were evaluated in each group. The kidney was placed inside an 18-mm NMR tube and the perfusion apparatus was positioned inside a wide-bore 9.4 T spectrometer. The kidney was perfused at 110 cm-H2O for 30 minutes while the spectrometer was shimmed on the 23Na signal. The flow rate and oxygen consumption were determined at 5 and 25 mins. After 30 mins, a solution of HP [1-13C]pyruvate dissolved in KH was injected into the kidney via the cannulated aorta and a series of 13C NMR spectra were acquired every 2 s with 20-deg pulses (total time = ~3 mins). The kidney was then perfused for additional 15 mins. The organ was freeze-clamped, pulverized, and extracted with 5-6% perchloric acid (PCA). The extract was neutralized, freeze-dried, and reconstituted in D2O/DSS/EDTA for high resolution 1H and 13C NMR analyses.Results and Discussion
Average renal oxygen consumptions of the kidneys perfused with pyruvate and pyruvate+octanoate was 12.9±3.9 and 11.5±2.5 µmol/min/g dw, respectively. The oxygen consumption difference between the two groups was not statistically different. Representative arrayed hyperpolarized 13C NMR spectra of kidneys receiving hyperpolarized [1-13C]pyruvate only or hyperpolarized [1-13C]pyruvate plus non-hyperpolarized octatanoate are shown in Fig. 1B. Summed spectra of these arrays are shown in Fig. 1C. From these data, rapid metabolism of 13C-pyruvate evident in both groups of kidneys as confirmed by the appearance of 13C-bicarbonate, 13C-alanine, and 13C-lactate. Lower 13C-bicarbonate production (~0.5-fold) was observed when octanoate was also present in the perfusate (Fig. 1D), demonstrating altered metabolism of pyruvate through pyruvate dehydrogenase enzyme as a result of octanoate present as a competing substrate for acetyl-CoA. Higher alanine and lactate signals were also observed in the kidneys receiving only 13C-pyruvate. Small peaks of 13C-malate were also observed in the pyruvate-only kidneys, but not detectable in the kidneys receiving pyruvate plus octanoate. These results indicate a more active metabolism of hyperpolarized 13C-pyruvate in the kidneys exposed to pyruvate as the only oxidizable substrate, as expected. A scheme showing the metabolism of [U-13C]pyruvate and [2,4,6,8-13C]octanoate and representative 1H-decoupled 13C NMR spectra of kidney extracts are shown in Fig. 2. 13C NMR isotopomer analyses of kidney extracts revealed that pyruvate and octanoate contributed equally (~40% each) to total acetyl-CoA while endogenous substrates such as triglycerides and glycogen accounted for the remainder of the acetyl-CoA source. When pyruvate was the only substrate, the majority of acetyl-CoA (~80%) was originated from pyruvate and the endogenous substrates contributed to the remainder of acetyl-CoA for energy production.Conclusions
We have demonstrated for the first time that real-time substrate metabolism can be evaluated in isolated perfused kidneys by hyperpolarized 13C NMR. Oxidative and reductive metabolism of pyruvate was detected in functioning kidneys and confirmed by isotopomer analyses of high-resolution 13C NMR of the kidney extracts. These results suggest that isolated perfused kidneys can serve as an excellent model to study renal metabolism by HP 13C NMR without the complexity of competing substrates present in vivo.Acknowledgements
This work supported by the NIH (5R37-HL034557 and 8P41-EB015908) and AHA 18POST34050049.References
- Moreno KX, et al. (2014) Real-time Detection of Hepatic Gluconeogenic and Glycogenolytic States Using Hyperpolarized [2-13C]Dihydroxyacetone. Journal of Biological Chemistry 289(52):35859-35867.
- Jin ES, Sherry AD, & Malloy CR (2016) An Oral Load of [13C3]Glycerol and Blood NMR Analysis Detect Fatty Acid Esterification, Pentose Phosphate Pathway, and Glycerol Metabolism through the Tricarboxylic Acid Cycle in Human Liver. Journal of Biological Chemistry 291(36):19031-19041.
- Wirthensohn G & Guder WG (1986) Renal substrate metabolism. Physiol Rev 66(2):469-497.
- Nishiitsutsuji-Uwo JM, Ross BD, & Krebs HA (1967) Metabolic activities of the isolated perfused rat kidney. Biochem J 103(3):852-862.
Figures
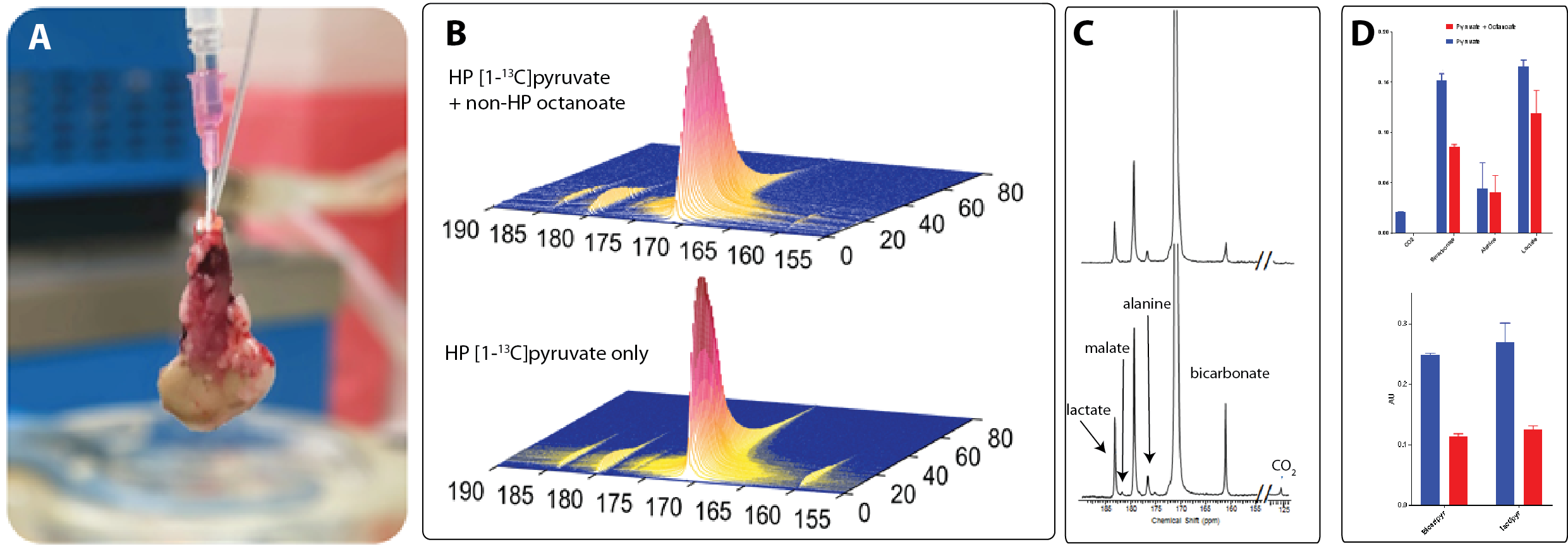
A) A picture of an isolated kidney cannulated and connected to a
perfusion apparatus; B) Arrayed 13C NMR spectra of kidneys receiving either hyperpolarized [1-13]pyruvate only (bottom) or hyperpolarized [1-13C]pyruvate
plus non-hyperpolarized octanoate (top); C) Summed spectra of the arrays shown
in B); and D) Bar graphs showing total area under the curve of all metabolites
normalized to the total 13C signals (top) and the lactate/pyruvate and
bicarbonate/pyruvate ratios from both groups of kidneys (bottom).
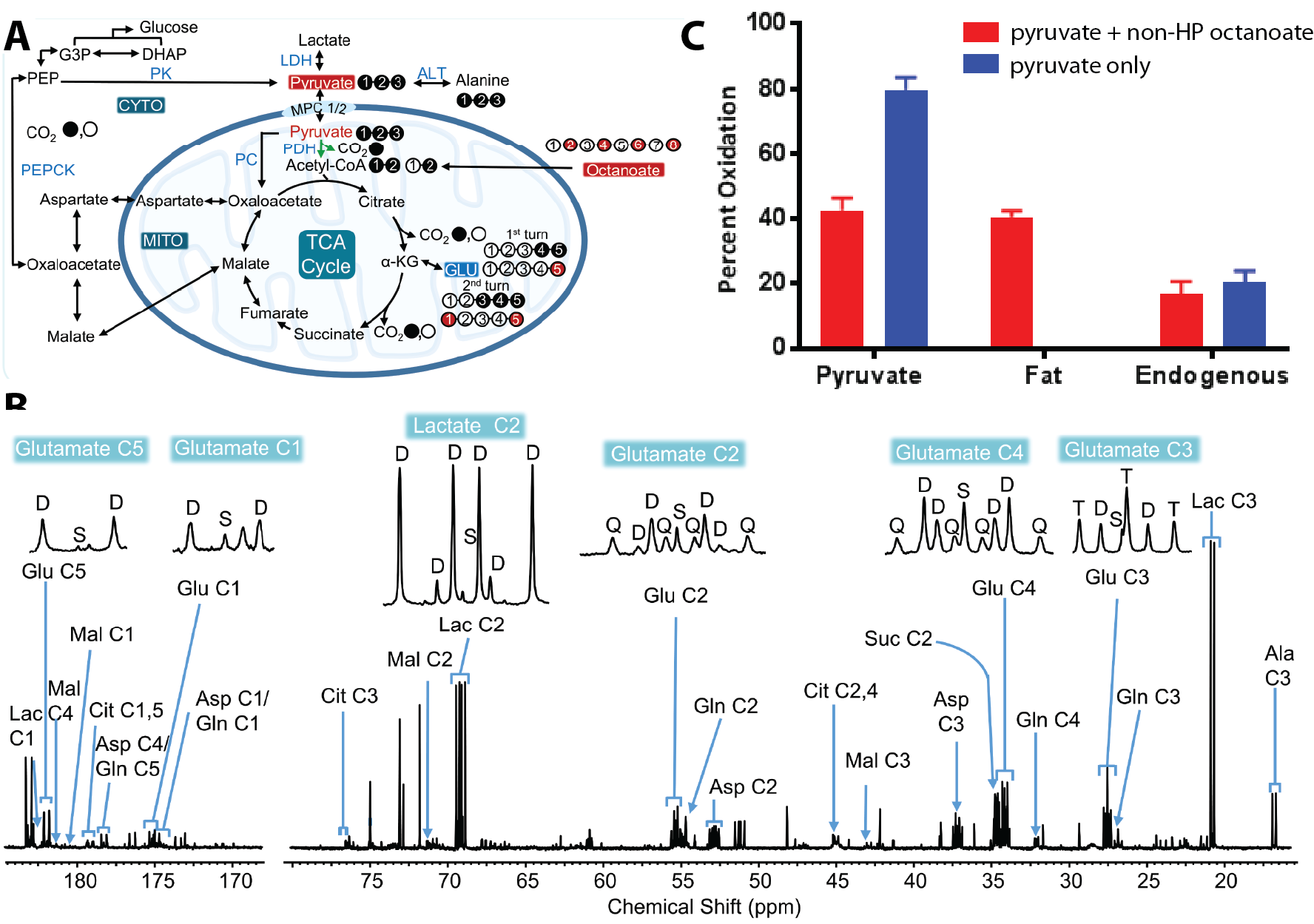
A) A metabolic scheme showing the metabolism of [U-13C]pyruvate
or [U-13C]pyruvate plus [2,4,6,8-13C]octanoate; B) Representative
high resolution 13C NMR spectra of tissue extract from a kidney perfused with [U-13C]pyruvate
plus [2,4,6,8-13C]octanoate; C) Bar graphs showing fractional
contributions of available oxidizable substrates to the total acetyl-CoA for
energy production.