4307
Pyruvate dehydrogenase kinase knockout restores hepatic pyruvate oxidation in STZ-induced diabetic mice as revealed by hyperpolarized 13C-MRI1Advanced Imaging Research Center, University of Texas Southwestern Medical Center, Dallas, TX, United States, 2Department of Biochemistry, University of Texas Southwestern Medical Center, Dallas, TX, United States, 3Department of Internal Medicine, University of Texas Southwestern Medical Center, Dallas, TX, United States, 4Department of Radiology, University of Texas Southwestern Medical Center, Dallas, TX, United States, 5Department of Chemistry, University of Texas at Dallas, Richardson, TX, United States
Synopsis
Pyruvate dehydrogenase (PDH) kinase (PDK) modulates mitochondrial oxidative metabolism of carbohydrate by phosphorylating and inhibiting PDH. PDK is, therefore, an important therapeutic target for treating and managing metabolic diseases. This study evaluated
Purpose
Pyruvate dehydrogenase (PDH) kinase (PDK) plays a critical role in regulating mitochondrial oxidative metabolism by inhibiting PDH via phosphorylation. Abnormalities of PDK activity are associated with a range of metabolic diseases including cancer and diabetes1. Drugs are under development to inhibit PDK activity and thereby stimulate carbohydrate oxidation. Therefore, the ability to assess in-vivo substrate metabolism as a result of PDK inhibition can be highly beneficial for understanding treatment and management of metabolic diseases2. Hyperpolarized 13C-MRI has been successfully utilized to detect PDH flux in humans. The aim of this study was to investigate in-vivo hepatic metabolism of hyperpolarized-13C pyruvate in PDK2/4 double knockout (DKO) and consequence of PDK2/4 double knockout (DKO) in streptozotocin (STZ) induced diabetic animals.Methods
Animals: Animal studies were approved by the Institutional Animal Care and Use Committee (IACUC). Metabolism of HP [1-13C]pyruvate was investigated in male C57BL/6J mice (WT) and PDK2/4 double knockout (DKO) mice in four groups: WT (n=3), DKO (n=3), STZ treated WT (n=2), and STZ treated DKO (n=2). To induce diabetes, the WT and DKO mice were fasted overnight and administered a single dose of 150 mg/kg STZ (freshly made in cold citrate buffer) intraperitoneally. After one week of STZ injection, a sample of blood was taken from the tail vein for blood glucose assessment. A jugular catheter was surgically implanted to facilitate injection for HP-13C MRS studies. Subsequently, animals were positioned in the MR scanner for metabolic evaluation using hyperpolarized [1‐13C]pyruvate.
Pyruvate polarization and injection: 24 µL [1-13C]-labeled pyruvic acid (Cambridge Isotope, USA) containing 15 mM trityl radical OX63 (GE Healthcare), and 2 mM Gd2+ was polarized at 1.4 K, 3.35 T for 1 h in a HyperSense® polarizer (Oxford Instruments). The polarized substrate was then quickly dissolved in PBS buffer (pH 7.4) and neutralized with NaOH. 200 µL of the hyperpolarized pyruvate (80 mM) was injected via the cannulated jugular vein catheter over 12–15 seconds.
In-vivo 13C MRSI: 13C spectra were acquired at 4.7 T MRI scanner (Agilent, Palo Alto, CA) using a Varian 60 mm dual tuned 1H/13C linear volume coil. Anatomical images were acquired using GEMS sequence to confirm the positioning (TE/TR, 4.0/80.5 ms; matrix size, 128 x 128; FOV, 35 x 35 mm; slice thickness, 2 mm; excitation flip angle, 20°). In-vivo 13C data acquisition was initiated immediately after the pyruvate injection with following parameters: TR: 2 s, flip angle: 10º, sweep width: 10 KHz, acquired points: 2048, frequency centered at 145 ppm. Blood plasma and livers were collected and freeze-clamped for metabolic measurements. Spectra were analyzed as described previously3. All data are presented as mean ± SEM.
Results and Discussions
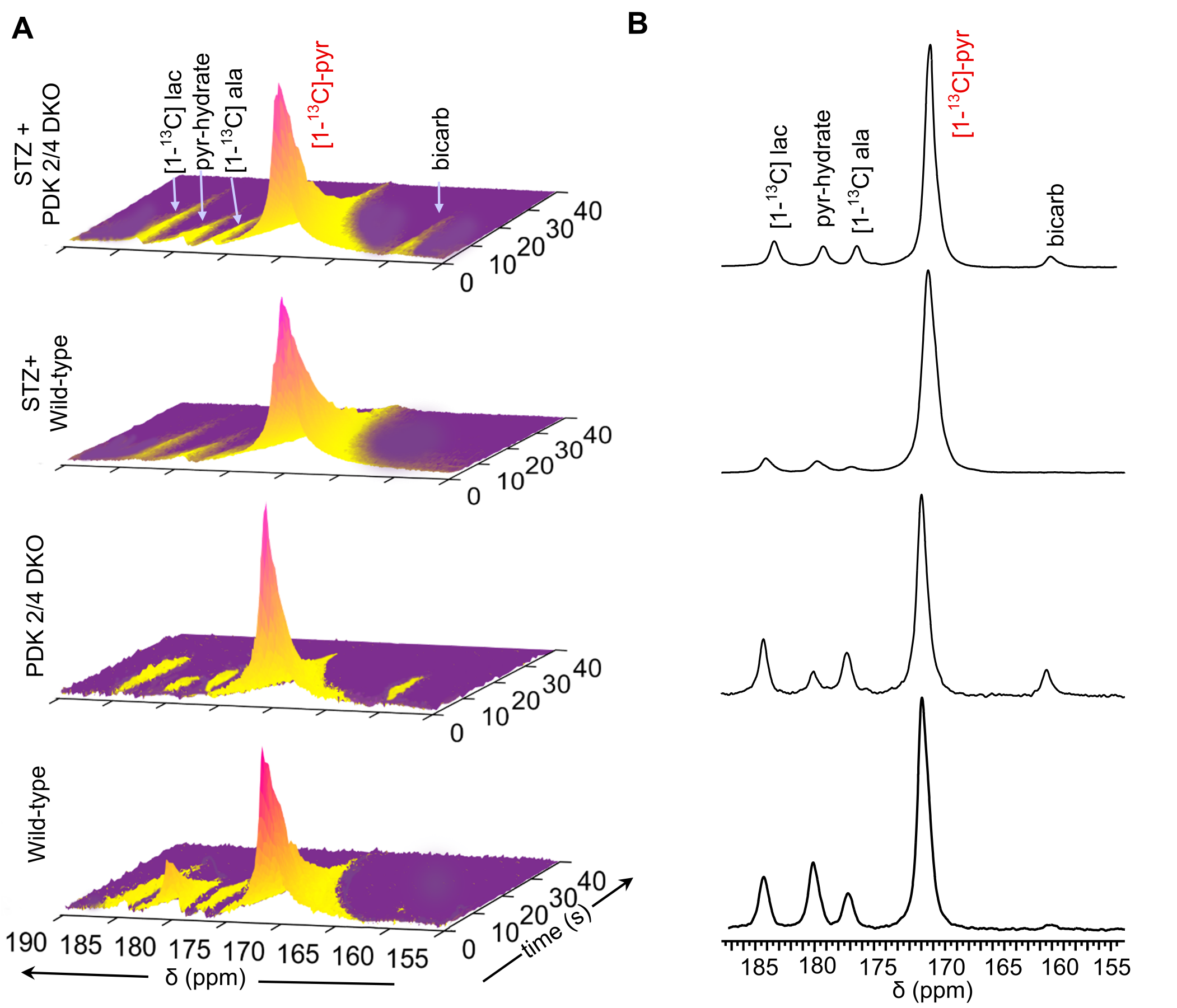
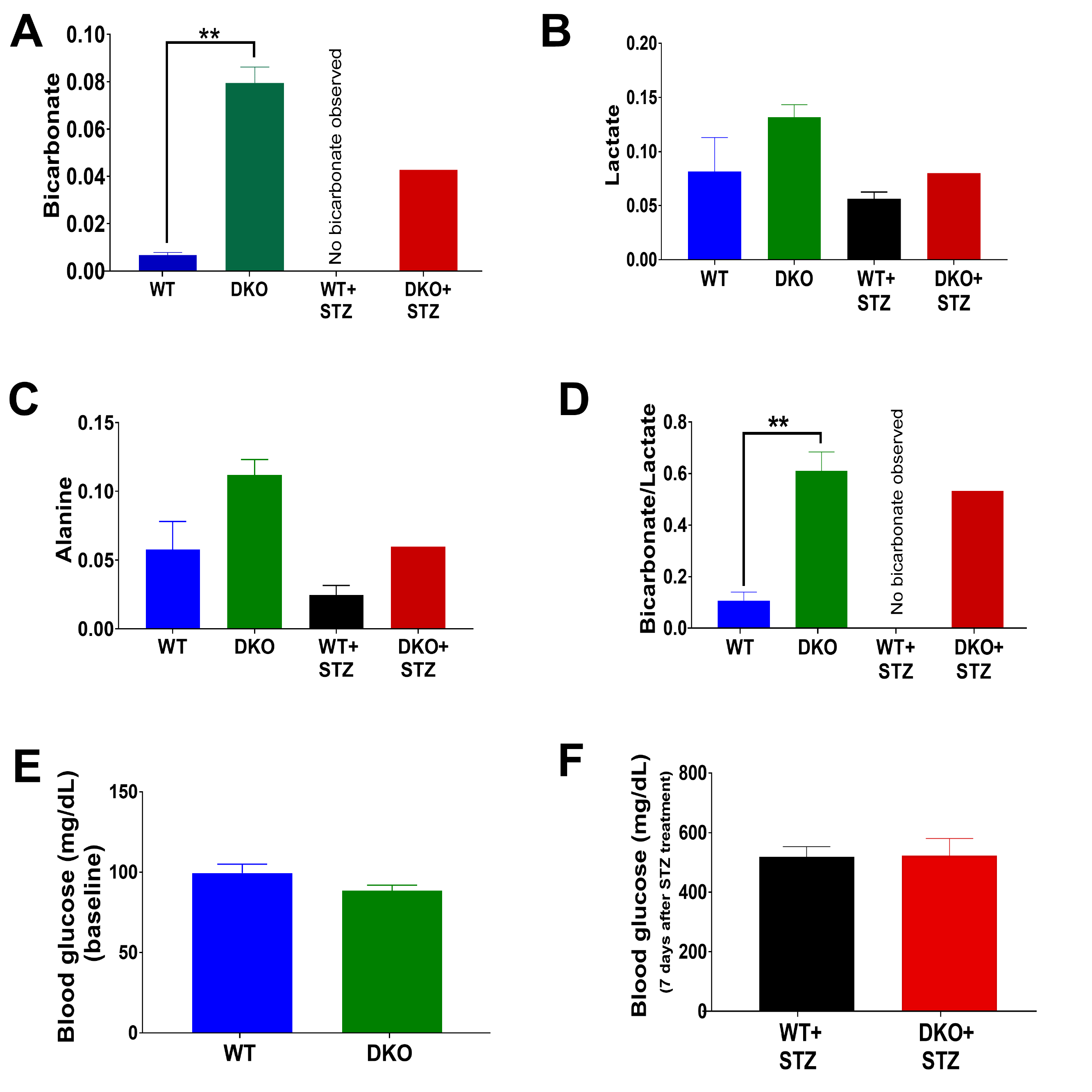
A scheme illustrating the metabolism of [1-13C]pyruvate via PDH and the role of PDK in pyruvate metabolism is shown in Figure 1A. Metabolism of pyruvate in the liver includes a reduction to lactate via LDH, transamination to alanine via ALT, carboxylation to oxaloacetate via PC and decarboxylation to acetyl-CoA via PDH. Figure 1B summarizes the study scheme of hepatic pyruvate metabolism assessment using hyperpolarized [1-13C]pyruvate. Representative sequential 13C MR spectra (Fig. 2A) acquired from mouse livers following an injection of 80-mM hyperpolarized [1-13C]pyruvate shows the dynamic changes in pyruvate metabolism. Summed 13C spectra (Fig. 2B) clearly show the production of [1-13C]lactate, [1-13C]alanine, and 13C-bicarbonate in the liver. Significantly increased bicarbonate production was observed in DKO mice. No detectable bicarbonate was observed in STZ-treated WT animals. While STZ-treatment results in decreased bicarbonate production in DKO animals, a significant degree of pyruvate oxidation was detected in these livers. A 43% decrease in bicarbonate signals was observed as a result of STZ-treatment of DKO mice (Fig. 3A). Reduced production of HP-[1-13C]lactate (Fig. 3B) and HP-[1-13C]alanine (Fig. 3C) were also observed in WT+STZ and DKO+STZ animals. A higher bicarbonate to lactate ratio in DKO mice emphasizes increased PDH flux, as expected, as a result of DKO in both normal and STZ-treated animals. Diabetic conditions of the STZ-treated mice were confirmed by high blood glucose levels (Fig. 3F). Circulating glucose concentrations were 518.5 and 522.5 mg/dL for WT+STZ and DKO+STZ, respectively. Baseline blood glucose levels (Fig. 3E) in WT and DKO were 99.5 and 88.5 mg/dL, respectively. Taken together, our results demonstrated that DKO of PDK2/4 stimulates pyruvate oxidation in both normal and type-1 diabetic mice.Conclusions
Using real-time hyperpolarized-13C MRI, we have demonstrated that STZ-induced type-1 diabetes significantly reduced oxidative pyruvate metabolism in the liver. The oxidative metabolism of pyruvate was partially restored by double knockout of PDKs as demonstrated by hyperpolarized 13C MRI. The results suggest that hyperpolarized-13C MRI could be used to evaluate the efficacy and therapeutic responses of metabolic drugs targeting PDK inhibition.Acknowledgements
This work supported by the NIH (5R37-HL034557 and 8P41-EB015908) and AHA 18POST34050049.References
1. Jeoung, N. H., Harris, C. R., and Harris, R. A. Regulation of pyruvate metabolism in metabolic-related diseases. Rev Endocr Metab Disord. 2014, 15, 99-110
2. Jeoung, N. H. Pyruvate Dehydrogenase Kinases: Therapeutic Targets for Diabetes and Cancers. Diabetes Metab J. 2015, 39, 188-197
3. Moreno, K. X., Moore, C. L., Burgess, S. C., Sherry, A. D., Malloy, C. R., and Merritt, M. E. Production of hyperpolarized (13)CO2 from [1-(13)C]pyruvate in perfused liver does reflect total anaplerosis but is not a reliable biomarker of glucose production. Metabolomics. 2015, 11, 1144-1156
Figures
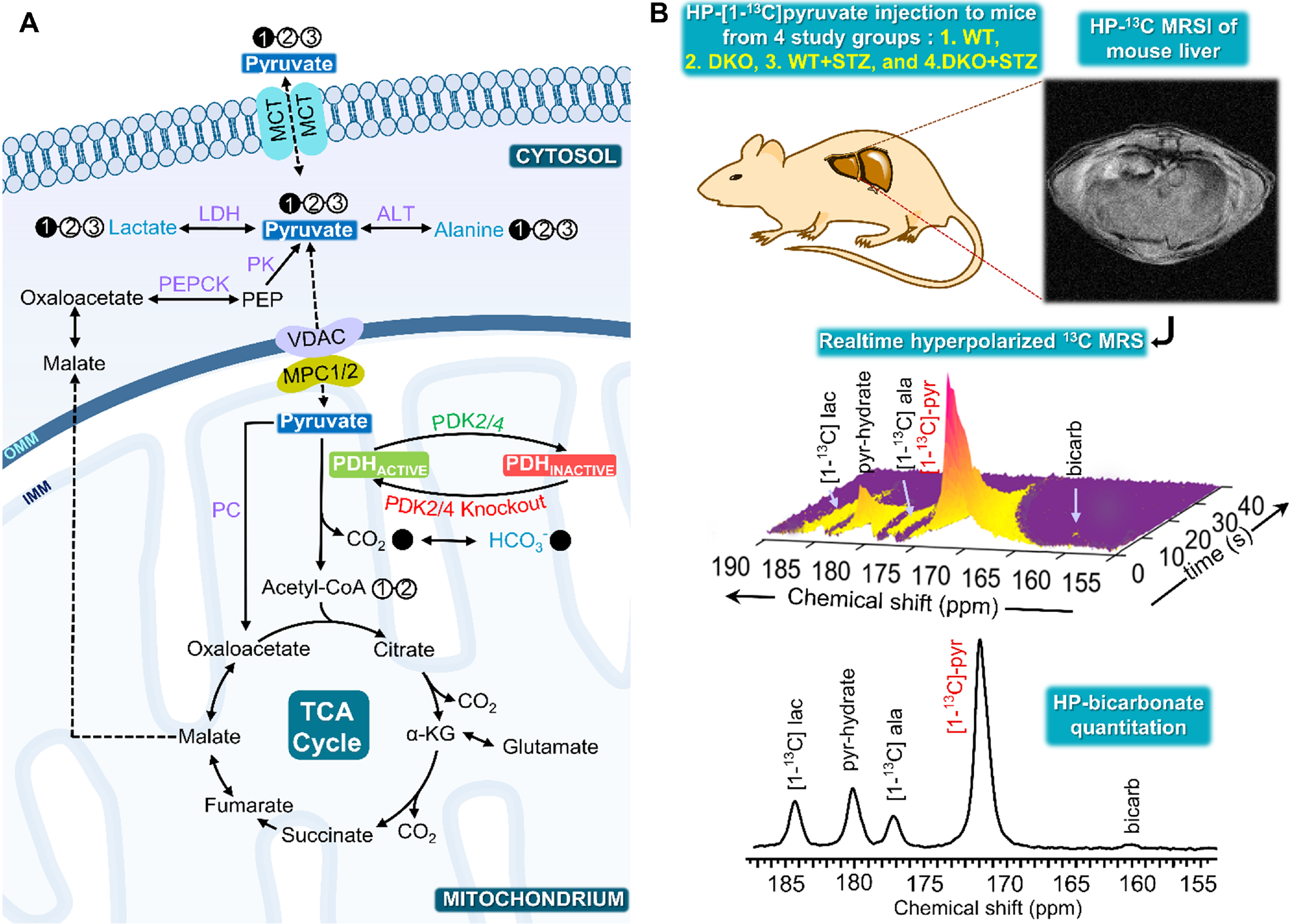
The filled circles designate 13C labeling and the empty circle represents 12C. The blue color metabolites detected in real-time hyperpolarized NMR following injection of HP [1-13C]pyruvate and the enzymes indicated in purple. ALT: Alanine transaminase; HP: Hyperpolarized; LDH: Lactate dehydrogenase; MCT: Monocarboxylate transporter; MPC1/2: Mitochondrial Pyruvate Carrier 1 and 2; PDC: Pyruvate dehydrogenase complex; PDK: Pyruvate dehydrogenase kinase; PC: Pyruvate carboxylase and PEPCK: Phosphoenolpyruvate carboxykinase.