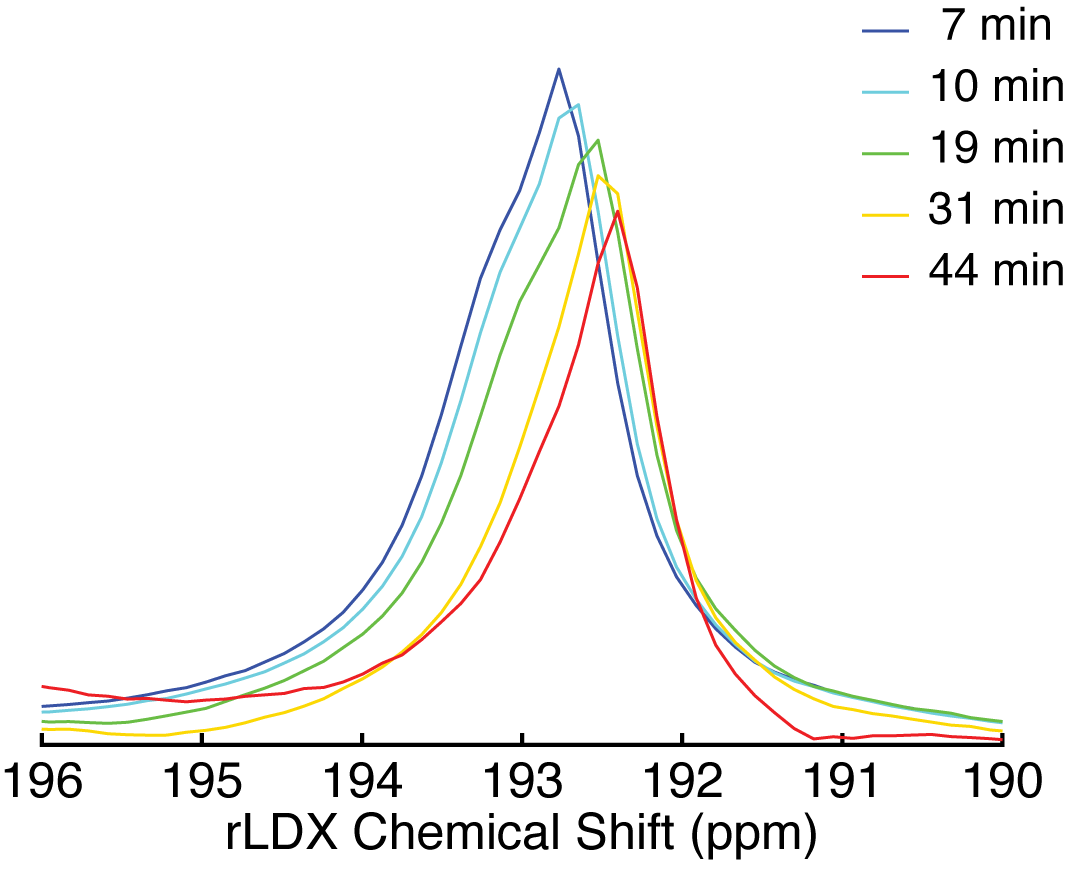4130
Absolute MR thermometry by hyperpolarized 129Xe in UCP1-/- animals reveals UCP1-independent thermogenesis1Physics and Astronomy, University of North Carolina at Chapel Hill, Chapel Hill, NC, United States, 2Biomedical Research Imaging Center, University of North Carolina at Chapel Hill, Chapel Hill, NC, United States
Synopsis
Direct and non-invasive temperature measurements of BAT in UCP1-/- mice could help to settle the debate on whether BAT in UCP1-/- mice is thermogenically competent and whether there exist other UCP1-independent mechanisms for non-shivering thermogenesis. Here we use hyperpolarized xenon MR thermometry for direct absolute temperature measurements of brown adipose tissue in UCP1-/- mice. Hyperpolarized xenon MR thermometry indicates a small but detectable temperature increase in the BAT of UCP1-/- mice after adrenergic stimulation of this tissue.
Introduction
Brown adipose tissue (BAT) is an organ specialized in heat production that has garnered much attention as a possible target for the treatment of obesity and diabetes1. For years the uncoupling protein one (UCP1) has been considered the hallmark of BAT thermogenesis, and the existence of other UCP1-independent thermogenic mechanisms has been a matter of debate for the past 20 years2–5. One critical tool for studying UCP1-independent mechanisms of thermogenesis has been the development of UCP1 knockout (KO) mice. However, in these animals BAT thermogenesis has been studied only indirectly, as methods to measure absolute temperature deep inside the body in very heterogeneous tissues like BAT have been lacking. While thermometry methods based on the temperature sensitivity of water proton chemical shift work extremely well in the brain, in anatomical regions with large magnetic susceptibility gradients6,7 these methods are generally prone to errors. Likewise, other 1H-based MR thermometry methods require pre-calibration of the temperature-sensitive parameters, which in vivo may be hard to obtain. Here we use a novel MR thermometry method based on the temperature sensitivity of the lipid-dissolved xenon chemical shift6,8,9 to assess whether the BAT of UCP1-/- animals is thermogenically competent.Methods
Localized 1H spectra and hyperpolarized 129Xe spectra were acquired from a region encompassing interscapular BAT, before and after norepinephrine (NE) injection, in anesthetized wild-type (WT) and UCP1 KO mice on a 9.4 T scanner (Bruker Biospin). During the entire experiment, bore temperature was regulated and maintained at 34°C by a forced-air heating system, while rectal temperature was monitored with an MR-compatible temperature probe. For the acquisition of hyperpolarized xenon spectra, mice were mechanically ventilated with a mixture of hyperpolarized 129Xe (polarized up to 18% by a commercial polarizer) and oxygen in a 75/25 volume ratio. From 1H and 129Xe spectra BAT absolute temperature values were then obtained as previously described10,11. Specifically, the temperature-sensitive frequency of xenon dissolved in lipids was referenced to the temperature insensitive frequency of nearby lipid protons to remove the effect of macro- and microscopic susceptibility gradients and obtain a referenced lipid-dissolved 129Xe (rLDX) chemical shift value from which absolute BAT temperature was derived9.Results
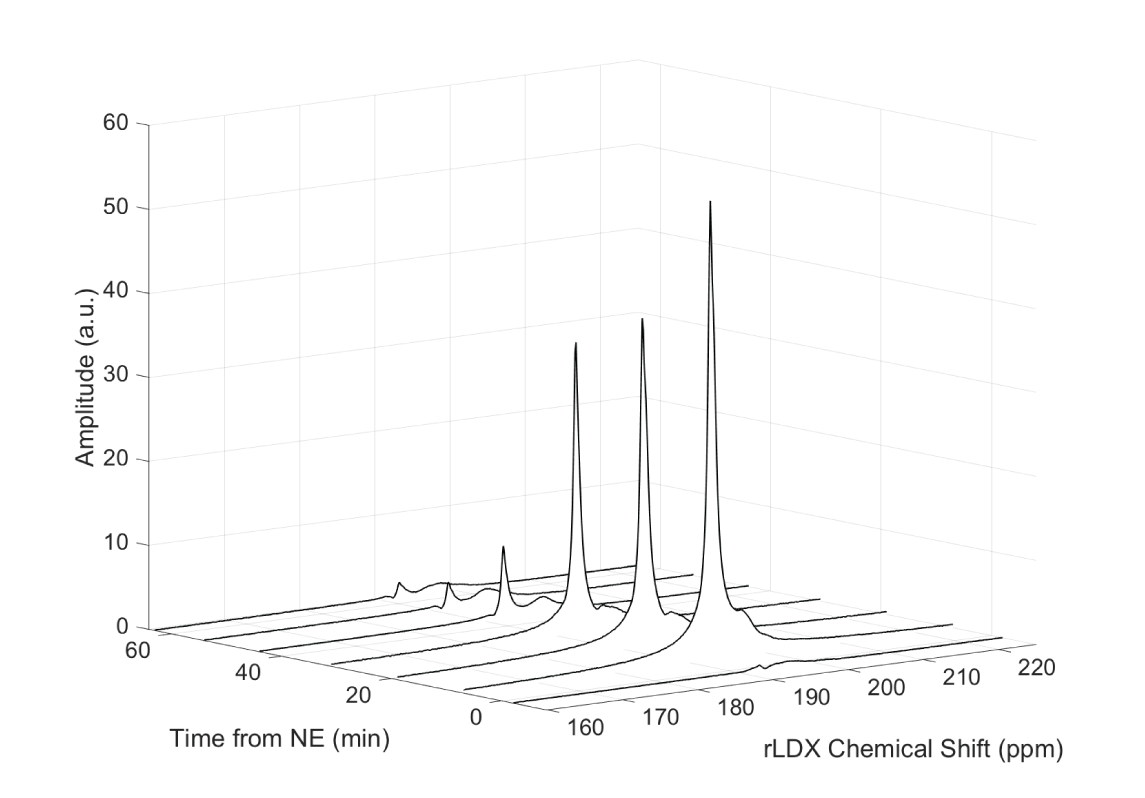
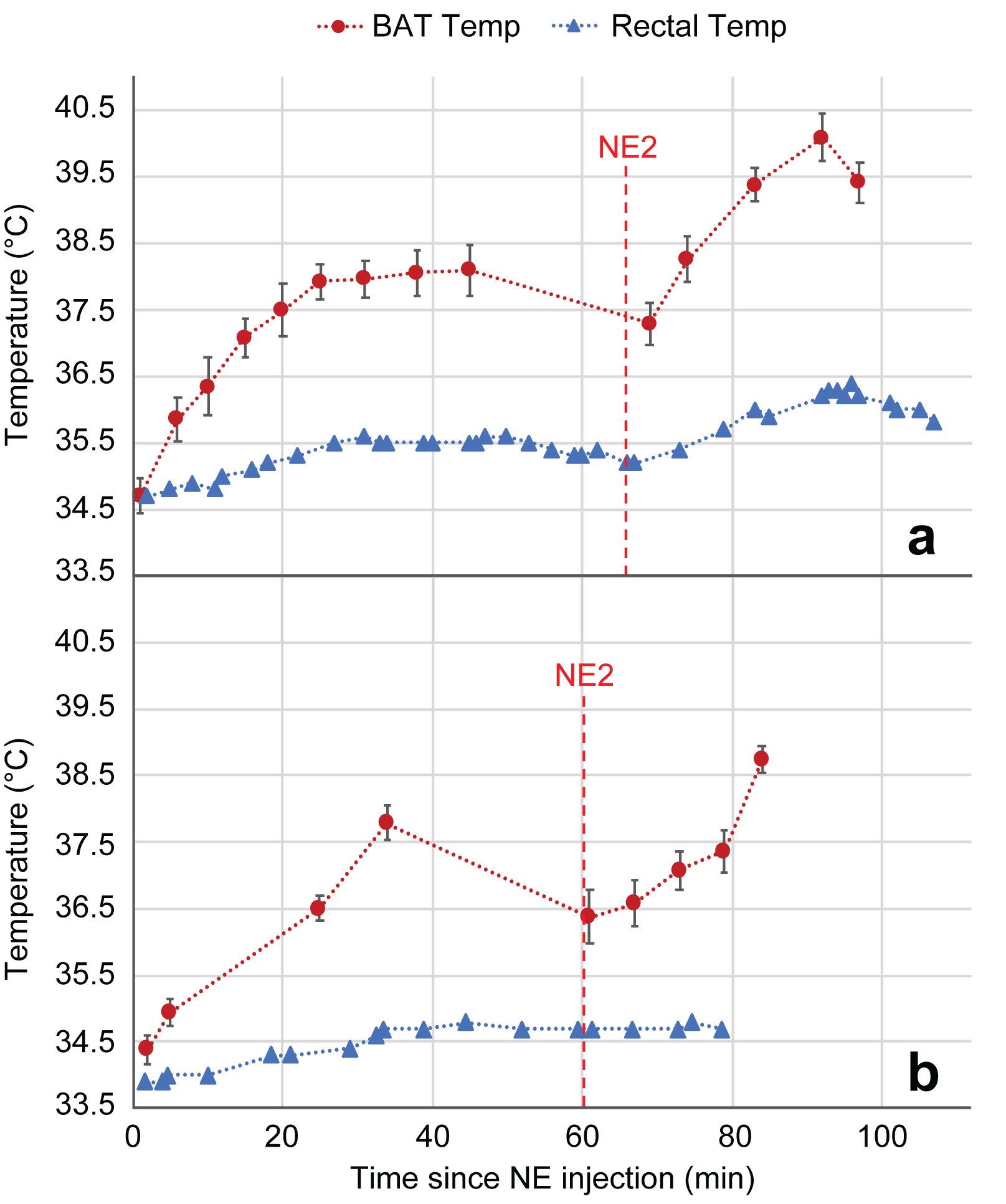
Figure 1 shows a waterfall plot of the rLDX peak before and during stimulation of BAT thermogenesis in a KO mouse. As in WT mice, stimulation of non-shivering thermogenesis leads to an increase in blood flow to BAT and to an increase in the rLDX signal intensity, which lasts until NE is degraded. Figure 2 shows the clear upfield drift of the rLDX chemical shift indicating an increase in BAT temperature following the first NE injection. Figure 3 shows examples of BAT and rectal temperature increase following two NE injections in a WT and a UCP1 KO mouse. In WT mice BAT stimulation leads to an overall increase in tissue temperature up to 5°C and to an increase in rectal temperature of almost 2°C. In KO mice the increase in BAT is smaller but still present.Discussion
The observed temperature increase in the adrenergically-stimulate BAT of KO mice shows that these animals are capable of recruiting some UCP1-independent pathway for thermogenesis in or very near to BAT. Rectal temperature changes only slightly and at a slower rate than BAT temperature, suggesting that the thermogenic mechanism responsible for the BAT and rectal temperature increase is located within the BAT. Since the rLDX chemical shift directly reports on the temperature of the lipid droplets contained in BAT in which xenon dissolves in relatively large quantities11,12, the method is a more sensitive and reliable way to directly measure cell temperature in brown adipocytes to answer biologically-relevant questions.Conclusion
The rLDX thermometry method serves as a sensitive and specific probe of absolute temperature in rodent BAT, possibly enabling studies of UCP1-independent non-shivering thermogenesis.Acknowledgements
This work was supported by NIH grant fund R01 DK108231.References
1. Nedergaard J, Cannon B. The Changed Metabolic World with Human Brown Adipose Tissue: Therapeutic Visions. Cell Metab. 2010;11:268–272. doi: 10.1016/j.cmet.2010.03.007.
2. Nedergaard J, Matthias A, Golozoubova V, Jacobsson A, Cannon B. UCP1: The original uncoupling protein - and perhaps the only one? J. Bioenerg. Biomembr. 1999;31:475–491.
3. Nedergaard J, Golozoubova V, Matthias A, Asadi A, Jacobsson A, Cannon B. UCP1: the only protein able to mediate adaptive non-shivering thermogenesis and metabolic inefficiency. Biochim. Biophys. Acta - Bioenerg. 2001;1504:82–106. doi: 10.1016/S0005-2728(00)00247-4.
4. Ukropec J, Anunciado RP, Ravussin Y, Hulver MW, Kozak LP. UCP1-independent Thermogenesis in White Adipose Tissue of Cold-acclimated Ucp1 -/- Mice. J. Biol. Chem. 2006;281:31894–31908. doi: 10.1074/jbc.M606114200.
5. Ikeda K, Kang Q, Yoneshiro T, et al. UCP1-independent signaling involving SERCA2b-mediated calcium cycling regulates beige fat thermogenesis and systemic glucose homeostasis. Nat. Med. 2017;23:1454–1465. doi: 10.1038/nm.4429.
6. Zhang L, Burant A, McCallister A, Zhao V, Koshlap KM, Degan S, Antonacci M, Branca RT. Accurate MR thermometry by hyperpolarized 129Xe. Magn. Reson. Med. 2017;78:1070–1079. doi: 10.1002/mrm.26506.
7. McCallister D, Zhang L, Burant A, Katz L, Branca RT. Effect of Microscopic Susceptibility Gradients on Chemical-Shift-Based Fat Fraction Quantification in Supraclavicular Fat. J. Magn. Reson. Imaging (online 4 October 2018) doi: 10.1002/jmri.26219.
8. Branca RT, He T, Zhang L, Floyd CS, Freeman M, White C, Burant A. Detection of brown adipose tissue and thermogenic activity in mice by hyperpolarized xenon MRI. Proc. Natl. Acad. Sci. U.S.A. 2014;111:18001–18006. doi: 10.1073/pnas.1403697111.
9. Antonacci MA, Zhang L, Degan S, Erdmann D, Branca RT. Calibration of methylene-referenced lipid-dissolved xenon frequency for absolute MR temperature measurements. Magn. Reson. Med. (online 14 September 2018) doi: 10.1002/mrm.27441.
10. Zhang L, Burant A, Mccallister A, Zhao V, Koshlap KM, Degan S, Antonacci M, Branca RT. Accurate MR thermometry by hyperpolarized 129Xe. Magn. Reson. Med. 2017;78:1070-1079. doi: 10.1002/mrm.26506.
11. Antonacci MA, Zhang L, Burant A, McCallister D, Branca RT. Simple and robust referencing system enables identification of dissolved-phase xenon spectral frequencies. Magn. Reson. Med. 2018;80:431–441. doi: 10.1002/mrm.27042.
12. Branca RT, McCallister A, Yuan H, et al. Accurate quantification of brown adipose tissue mass by xenon-enhanced computed tomography. Proc. Natl. Acad. Sci. 2018;115:174–179. doi: 10.1073/pnas.1714431115.
Figures