3752
Optogenetically-evoked spindle-like activity from thalamus propagates brain-wide and enhances rsfMRI connectivity1Laboratory of Biomedical Imaging and Signal Processing, The University of Hong Kong, Hong Kong, China, 2Department of Electrical and Electronic Engineering, The University of Hong Kong, Hong Kong, China
Synopsis
Hierarchical nesting of spindles and slow oscillations have been identified as a unique characteristic for long-range functional neural integrations in
Purpose
Spontaneous oscillatory neural activities, such as slow oscillations and spindles, is a fundamental characteristic of long-range functional neural integrations in large-scale brain networks1-3. In particular, thalamo-cortical spindles (7-14 Hz activity lasting 0.5~3s) actively communicate with multiple high-order systems such as the limbic system (e.g., hippocampus and cingulate cortex), basal ganglia and sensory cortices to subserve numerous cognitive functions2,4-7. Despite its importance, our understanding of the large-scale distribution and functions of thalamo-cortical spindles remains poor. This is due to the lack of suitable neurotechnologies that can both initiate specific neural events (e.g., spindles), and monitor their activities and functions in brain-wide networks. Recently, resting-state fMRI (rsfMRI) has emerged as a valuable tool to not only map large-scale and long-range networks8,9, but increasingly used in the clinical setting. However, rsfMRI only reflects the gross outcome of complex cascaded neural activities of various brain networks, leaving many important characteristics of the identified networks unresolved. Hence, it would be extremely valuable if the underlying neural bases of rsfMRI connectivity/networks could be investigated with the ability to initiate functionally-relevant large-scale neural activity such as thalamo-cortical spindles. In this study, we develop an optogenetic fMRI platform to induce large-scale spindle-like activities through the stimulation of the ventral posteromedial thalamus (VPM) excitatory neurons and monitor their influences on brain-wide rsfMRI connectivity.Method
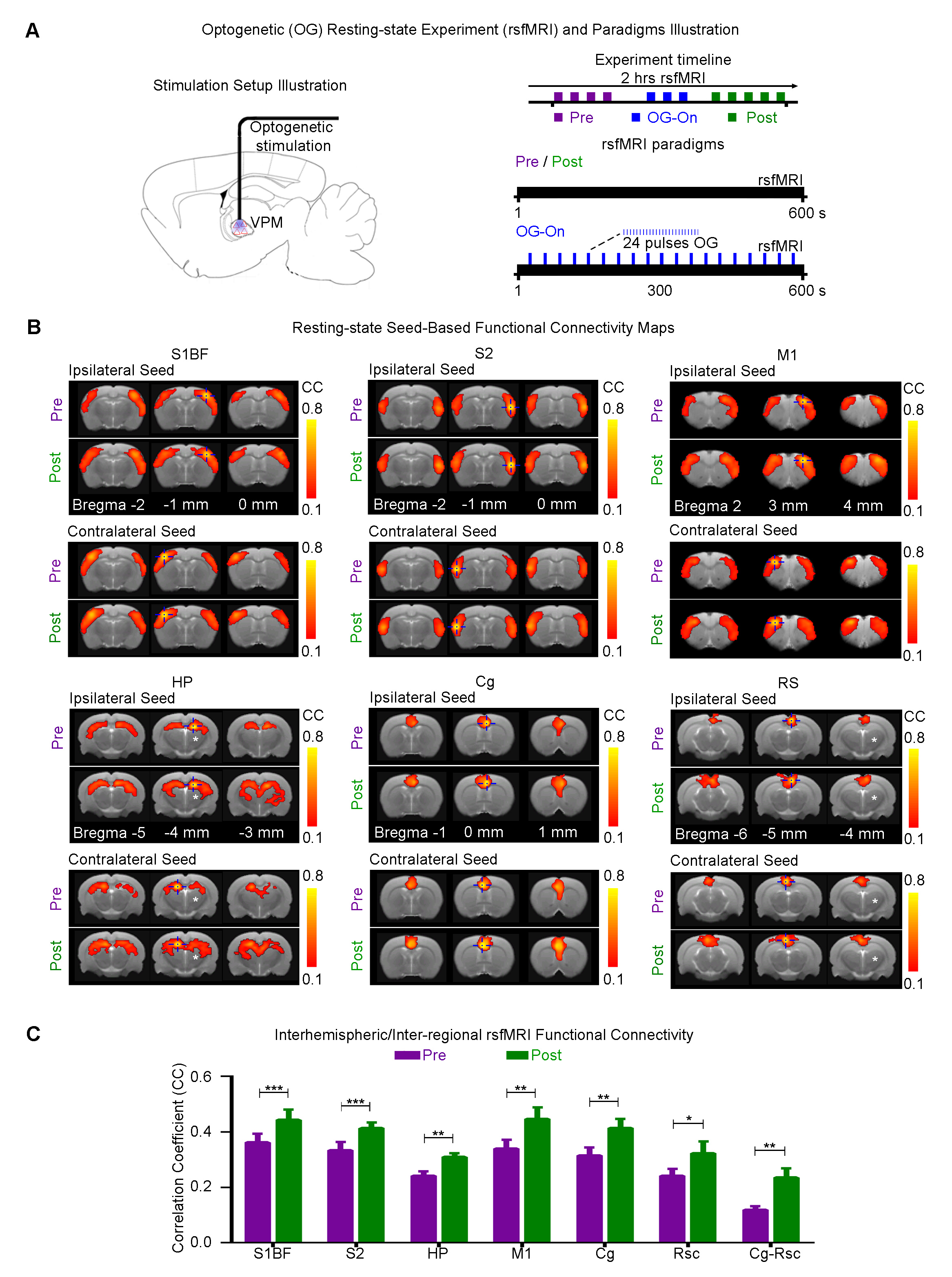
Animal preparation and optogenetic stimulation: 3μl AAV5-CaMKIIα::ChR2(H134R)-mCherry was injected to VPM of SD rats (n=12, 200-250g, male). Four weeks after injection, an opaque optical fiber cannula was implanted at the injection site. All experiments were performed under 1.0% isoflurane. 4 and 24 blue (473nm) light pulses (0.5 and 3s duration) at 8Hz were presented every 30s (10% duty cycle, 40mW/mm2; Figure 1).
Optogenetic fMRI, rsfMRI and electrophysiology experiments: MRI data was acquired at 7T using GE-EPI. Coherence analysis10 was applied to identify significant BOLD responses. Seed-based analysis was applied to map and quantify interhemispheric rsfMRI connectivity of primary somatosensory barrel field (S1BF), secondary somatosensory (S2), cingulate (Cg), retrosplenial (RS) cortices, and hippocampus (HP). Electrophysiology data (n=3) was acquired using multi-depth electrodes (16 channels) and the computed current source density was used to analyze the evoked primary and secondary slow oscillatory responses.
Results
Brain-wide fMRI mapping of spindle-like optogenetic stimulation: We detected robust positive BOLD activation in the sensorimotor cortices, thalamus and midbrain regions, higher-order sensory and motor-related cortices, limbic regions, and basal ganglia upon 24 pulses 8Hz optogenetic stimulation of VPM excitatory neurons (Figure 2). Responses include . Additionally, we observed similar brain-wide BOLD activations when the number of stimulation pulses was decreased to 4, albeit with lower response amplitudes (Figure 3).
Optogenetically-evoked spindle-like activity increases brain-wide cortical rsfMRI connectivity, including higher order regions such as prefrontal cortices and hippocampus: We found that after (post) 24 pulses VPM stimulation, the strength of interhemispheric connectivity increased significantly in S1BF, S2, HP, Cg, RS and M1 (Figure 4A, B). Interestingly, we observed that the connectivity between Cg and RS are significantly enhanced. We also observed an increase in the spatial extent of all connectivity maps.
Characteristics of optogenetically evoked spindle-like neural activity in ipsilateral S1: We detected evoked waxing-and-waning shaped spindle-like local field potentials (LFPs) during 4/24 pulses stimulations in the primary somatosensory cortex (S1) (Figure 5B, C). The waxing and waning phases short-tailed by their progressive increase and subsequent decrease of evoked LFP amplitudes. We also observed slow secondary neural oscillatory responses after the primary evoked LFP.
Discussion
Our results demonstrate that spindle-like optogenetic stimulation of the VPM thalamocortical excitatory neurons activated brain-wide regions including sensory- and motor-related thalamo-cortical and midbrain regions, limbic system, and basal-ganglia. Spindle-related BOLD activity was robustly evoked across multiple deep and small structures in limbic system (e.g., amygdala, entorhinal areas) and basal ganglia, which are difficult to be delineated in previous human fMRI and electrophysiology studies. Compared to the ipsilateral response bias in the subcortical regions, which are of low amplitudes and with short-tailed BOLD responses, the cortices have widespread, large amplitude and long-tailed BOLD responses. These observations corroborated with a previous report showing that the cortex amplifies and sustains subcortical spindles11. Furthermore, we demonstrated that thalamo-cortical spindle-like optogenetic stimulation significantly enhanced interhemispheric rsfMRI connectivity in S1BF, S2, HP, Cg, RS and M1 and interregional rsfMRI connectivity between Cg and RS. Our electrophysiology results verified the thalamically-evoked spindle-like activity in S1BF with similar characteristics as those reported for spontaneous spindles12,13 and optogenetically evoked spindle-like activity14,15. In addition, the slow oscillatory responses to the spindle-like activity suggests that the interaction between spindle-like activity and slow oscillations may participate in supporting brain-wide rsfMRI connectivity. In conclusion, our study demonstrate for the first time that functionally-relevant thalamo-cortical spindle-like neural activities can contribute to brain-wide rsfMRI connectivity.Acknowledgements
This work was supported by the Hong Kong Research Grant Council (C7048-16G and HKU17103015 to E.X.W.).References
1 Logothetis, N. K. Neural-Event-Triggered fMRI of large-scale neural networks. Curr Opin Neurobiol 31, 214-222, doi:10.1016/j.conb.2014.11.009 (2015).
2 Staresina, B. P. et al. Hierarchical nesting of slow oscillations, spindles and ripples in the human hippocampus during sleep. Nat Neurosci 18, 1679-1686, doi:10.1038/nn.4119 (2015).
3 Florin, E., Watanabe, M. & Logothetis, N. K. The role of sub-second neural events in spontaneous brain activity. Curr Opin Neurobiol 32, 24-30, doi:10.1016/j.conb.2014.10.006 (2015).
4 Schabus, M. et al. Hemodynamic cerebral correlates of sleep spindles during human non-rapid eye movement sleep. Proc Natl Acad Sci U S A 104, 13164-13169, doi:10.1073/pnas.0703084104 (2007).
5 Mak-McCully, R. A. et al. Coordination of cortical and thalamic activity during non-REM sleep in humans. Nat Commun 8, 15499, doi:10.1038/ncomms15499 (2017).
6 Berke, J. D., Okatan, M., Skurski, J. & Eichenbaum, H. B. Oscillatory entrainment of striatal neurons in freely moving rats. Neuron 43, 883-896, doi:10.1016/j.neuron.2004.08.035 (2004).
7 Saunders, A. et al. A direct GABAergic output from the basal ganglia to frontal cortex. Nature 521, 85-89, doi:10.1038/nature14179 (2015).
8 Gusnard, D. A., Raichle, M. E. & Raichle, M. E. Searching for a baseline: functional imaging and the resting human brain. Nat Rev Neurosci 2, 685-694, doi:10.1038/35094500 (2001).
9 Bullmore, E. & Sporns, O. Complex brain networks: graph theoretical analysis of structural and functional systems. Nat Rev Neurosci 10, 186-198, doi:10.1038/nrn2575 (2009).
10 Lee, J. H. et al. Global and local fMRI signals driven by neurons defined optogenetically by type and wiring. Nature 465, 788-792, doi:10.1038/nature09108 (2010).
11Beenhakker, M. P. & Huguenard, J. R. Neurons that fire together also conspire together: is normal sleep circuitry hijacked to generate epilepsy? Neuron 62, 612-632, doi:10.1016/j.neuron.2009.05.015 (2009).
12 Coppieters 't Wallant, D., Maquet, P. & Phillips, C. Sleep Spindles as an Electrographic Element: Description and Automatic Detection Methods. Neural Plast 2016, 6783812, doi:10.1155/2016/6783812 (2016).
13 Contreras, D. & Steriade, M. Spindle oscillation in cats: the role of corticothalamic feedback in a thalamically generated rhythm. The Journal of Physiology 490, 159-179 (1996).
14 Kim, A. et al. Optogenetically induced sleep spindle rhythms alter sleep architectures in mice. Proc Natl Acad Sci U S A 109, 20673-20678, doi:10.1073/pnas.1217897109 (2012).
15 Latchoumane, C.-F., Ngo, H.-V., Born, J. & Shin, H.-S. Thalamic Spindles Promote Memory Formation during Sleep through Triple Phase-Locking of Cortical, Thalamic, and Hippocampal Rhythms. Neuron 95, 424-435, doi:10.1016/j.neuron.2017.06.025 (2017).
Figures