3730
Sensorineural hearing loss and cognitive impairments : contributions of thalamus using multimodal MR1Department of Radiology, Jiangsu Key laboratory of Molecular Imaging and Functional Imaging, Nanjing, China, 2Department of Radiology, Zhongda Hospital, Nanjing, China, 3Medical School of Southeast Univeristy, Nanjing, China
Synopsis
In present study, we systematically investigated the structure, function and perfusion features of the TH and its seven subdivisions through multimodal MRI, aiming to uncover the potential role of the TH in SNHL and its correlations with cognition measurements.
Introduction
Sensorineural hearing loss (SNHL) is accounting for 90% of reported hearing loss (HL) (1), leading to difficulties in daily life (2). Cross-section and longitudinal evidence suggested that auditory system dysfunction, including age-related HL and congenital deafness, contributed to cognitive impairments (3-5), which is a result of neural plasticity. However, so far, the underlying mechanism of reduced audibility induced negative plasticity need to be further explored. As a paired deep structure in brain, the thalamus (TH) is traditionally conceptualized as a sensory hub (6). But it is still unclear whether the TH is the potential factor for SNHL-related cognitive impairments.Methods
We recruited thirty-seven subjects with long term bilateral SNHL and 38 healthy controls. All subjects underwent the pure tone audiometry (PTA) test at six frequencies (0.25, 0.5, 1, 2, 4, 8 kHz) and series of cognition tests, including Mini-mental state examination (MMSE), symbol digit modalities test (SDMT) and auditory verbal learning test (AVLT). After collection of structural and functional data, only forty-five subjects completed the ASL sequence because of long scanning time.
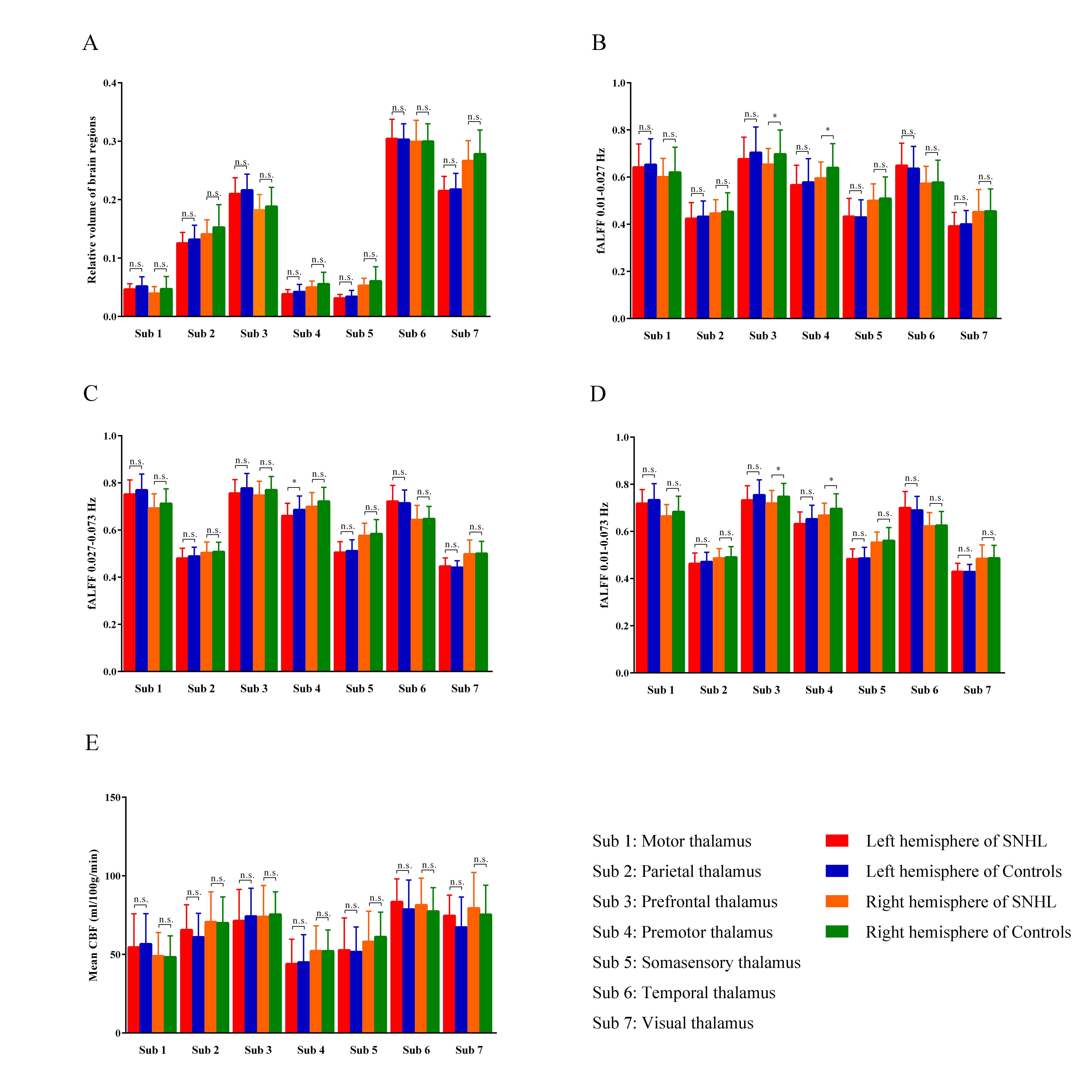
The TH was defined according to Anatomical Automatic Labeling (AAL) atlas, then divided into seven subdivisions according to SPM Anatomy toolbox, including motor (sub 1), parietal (sub 2), prefrontal (sub 3), premotor (sub4), somatosensory (sub 5), temporal (sub 6) and visual (sub 7) regions (see Figure 2). These ROIs were applied to all individual multimodal maps by normalizing to different template space. For left, right TH and subdivisions of participant, we quantified 5 measurements: (a) relative volume; (b) fALFF value at 0.01-0.027 Hz; (c) fALFF value at 0.027-0.073 Hz; (d) fALFF value at 0.01-0.073 Hz; (e) mean CBF. Moreover, we further explored the connectivity between TH subdivision and other brain area.
Results
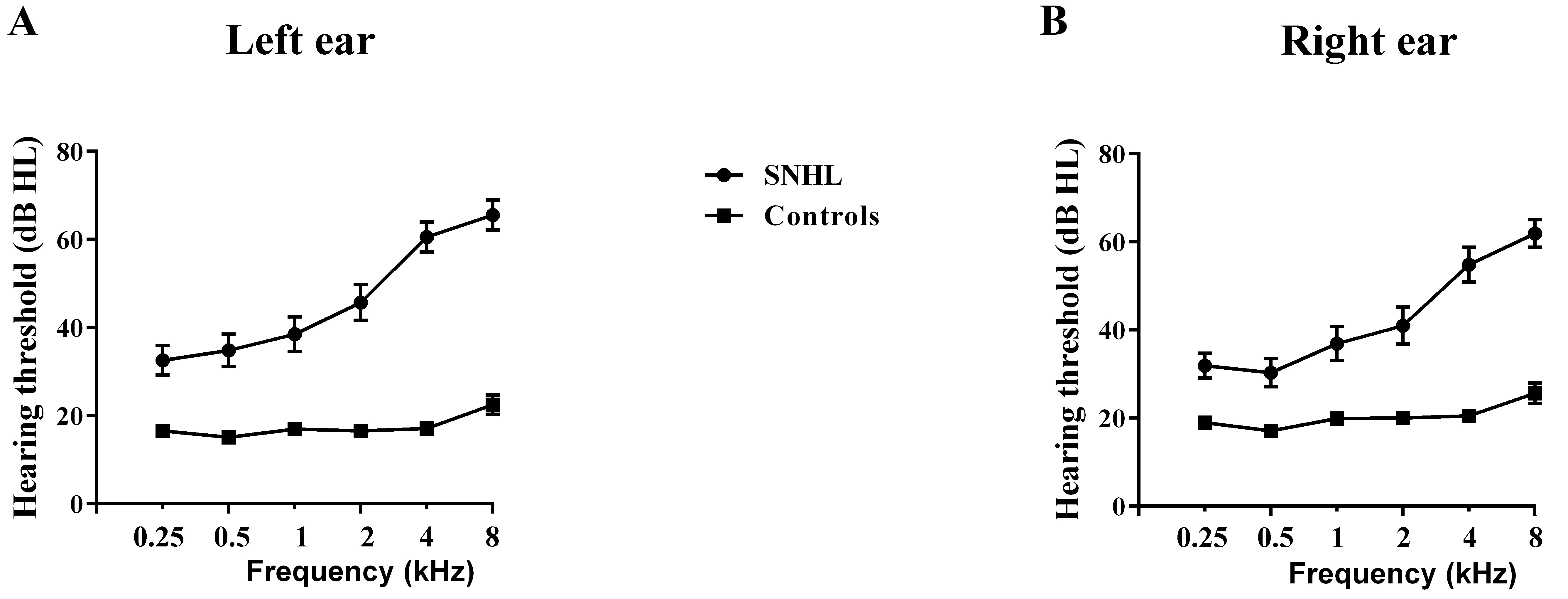
The diagnosis of SNHL was determined by PTA test (Figure 1). In terms of structure, we observed no significant group effects in bilateral TH and 7 subregions (Figure 2A).
The fALFF results of left and right TH showed no difference in slow 5 (0.01-0.027), slow 4 (0.027-0.073) and combined band (0.01-0.073). fALFF values in right prefrontal TH and premotor TH were found to be significant in slow 5 (Figure 2B). Only left premotor TH was of weak significance in slow 4 (p = 0.049, Figure 2C). In the combined band, right prefrontal and premotor were displayed to be prominently different (Figure 2D). No significance was observed in perfusion too (Figure 2E).
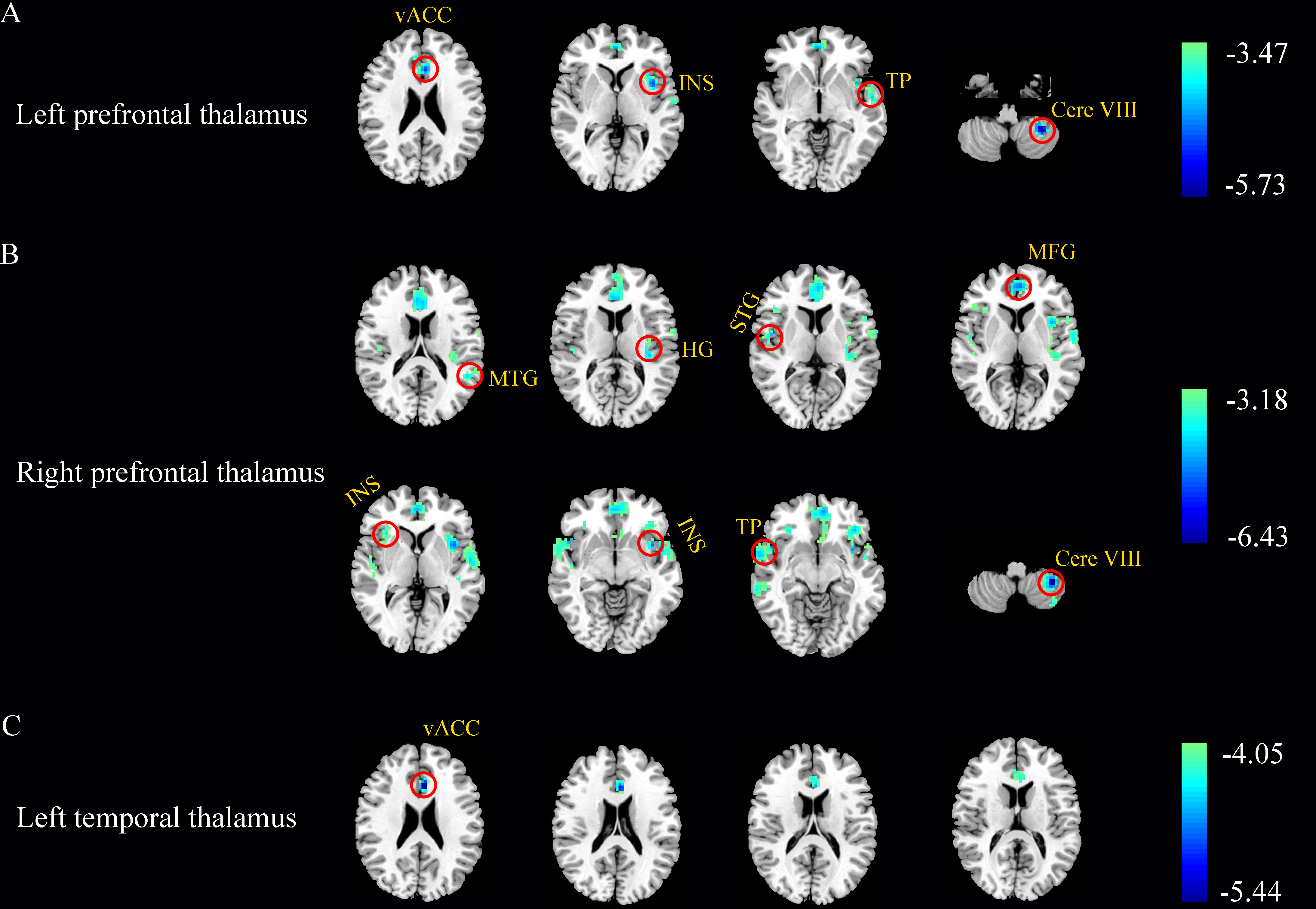
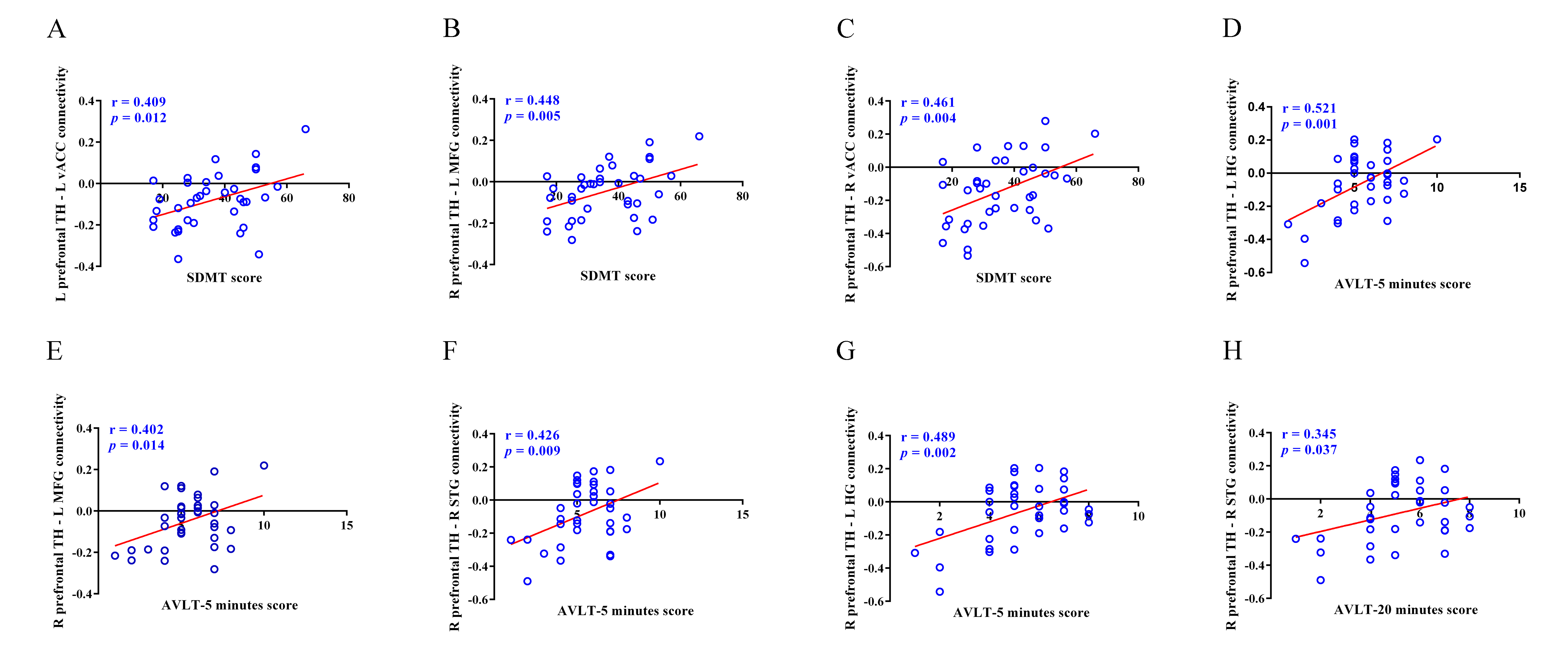
After FDR correction (p < 0.05), left prefrontal TH showed weakened connectivity with left cerebellum lobe VIII, ventral anterior cingulate cortex (vACC), insula and superior temporal gyrus (STG) in SNHL group. The voxel-wise analysis revealed group differences in right prefrontal TH connectivity with left cerebellum lobe VIII, insula, medial frontal gyrus (MFG), Heschl’s gyrus (HG) and media temporal gyrus (MTG), as well as right temporal pole (TP), STG and insula. Meanwhile, left temporal TH and left vACC connectivity was also decreased in SNHL comparing to Controls (Figure 3). Person’s correlation analysis was calculated between findings related with TH subregions and neuropsychological tests. And some of functional connectivity were positively correlated with cognitive tests (Figure 4).
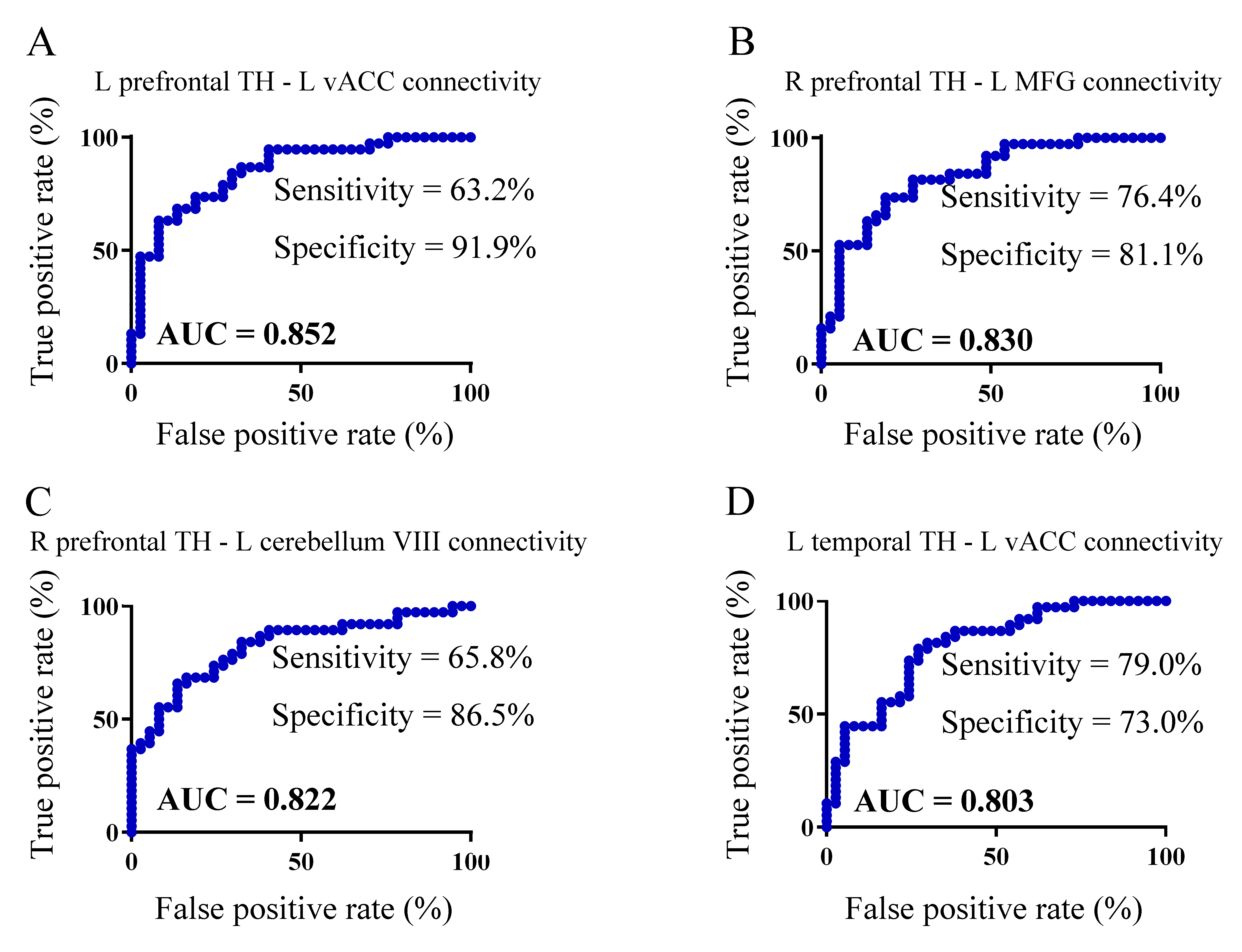
ROC analysis revealed that four connectivity characteristics of TH subregions (e.g. left prefrontal TH-left vACC FC, right prefrontal TH-left MFG FC, right prefrontal-left cerebellum VIII FC and left temporal TH-left vACC FC) showed better performance (0.8 < AUC < 1) in the classification between SNHL and Control groups, comparing with other measurements (Figure 5).
Discussion
Normal sensory input is the guarantee of the integrity of TH and brain functioning. Microstructure MRI found increased mean diffusivity and radial diffusivity in subjects with tinnitus (7). Increased volume of the TH (8) and downregulation of the inhibitory neurotransmitter (9) was also indicated in chronic tinnitus, but few study shed light on the role of TH in acquired SNHL. Moreover, the TH not only relays information from sensory organs or subcortical structures to the cortex, but also regulates the flow of information between cortical regions (10, 11). Emerging evidence has linked the TH to cognition via thalamocortical radiations, suggesting that the TH is the key structure in cognition processing, including memory, attention and executive control (12-14). Our study provides direct and comprehensive evidence for the involvement of TH in SNHL subjects, suggesting a potential therapeutic target in SNHL-associated cognitive impairments.Conclusion
We firstly identified the crucial role of TH by treating it as a single structure and seven distinct subregions using multimodal imaging approaches, underlying the potential mechanism of TH-related cognitive impairments in SNHL.Acknowledgements
We thank Xu Feng and Zhi-Chun Huang, E,N.T.department, Affiliated Zhongda Hospital of Southeast University, for their assistance with data collection. We also thank Jing Zhao, E,N.T.department, Affiliated Zhongda Hospital of Southeast University, for her assistance with auditory evaluation.References
1. Li L, Chao T, Brant J, O'Malley B, Jr., Tsourkas A, Li D. Advances in nano-based inner ear delivery systems for the treatment of sensorineural hearing loss. Advanced drug delivery reviews. 2017;108:2-12. 2. Dai L, Best V, Shinn-Cunningham BG. Sensorineural hearing loss degrades behavioral and physiological measures of human spatial selective auditory attention. Proceedings of the National Academy of Sciences of the United States of America. 2018;115(14):E3286-E95.
3. Heywood R, Gao Q, Nyunt MSZ, Feng L, Chong MS, Lim WS, et al. Hearing Loss and Risk of Mild Cognitive Impairment and Dementia: Findings from the Singapore Longitudinal Ageing Study. Dementia and geriatric cognitive disorders. 2017;43(5-6):259-68.
4. Swords GM, Nguyen LT, Mudar RA, Llano DA. Auditory system dysfunction in Alzheimer disease and its prodromal states: A review. Ageing research reviews. 2018;44:49-59.
5. Rutherford BR, Brewster K, Golub JS, Kim AH, Roose SP. Sensation and Psychiatry: Linking Age-Related Hearing Loss to Late-Life Depression and Cognitive Decline. The American journal of psychiatry. 2018;175(3):215-24.
6. Lin F, Zivadinov R, Hagemeier J, Weinstock-Guttman B, Vaughn C, Gandhi S, et al. Altered nuclei-specific thalamic functional connectivity patterns in multiple sclerosis and their associations with fatigue and cognition. Multiple sclerosis. 2018:1352458518788218.
7. Benson RR, Gattu R, Cacace AT. Left hemisphere fractional anisotropy increase in noise-induced tinnitus: A diffusion tensor imaging (DTI) study of white matter tracts in the brain. Hearing Res. 2014;309:8-16.
8. Muhlau M, Rauschecker JP, Oestreicher E, Gaser C, Rottinger M, Wohlschlager AM, et al. Structural brain changes in tinnitus. Cerebral cortex. 2006;16(9):1283-8.
9. Brozoski T, Odintsov B, Bauer C. Gamma-aminobutyric acid and glutamic acid levels in the auditory pathway of rats with chronic tinnitus: a direct determination using high resolution point-resolved proton magnetic resonance spectroscopy (H-MRS). Frontiers in systems neuroscience. 2012;6:9.
10. Hwang K, Bertolero MA, Liu WB, D'Esposito M. The Human Thalamus Is an Integrative Hub for Functional Brain Networks. J Neurosci. 2017;37(23):5594-607.
11. Herrero MT, Barcia C, Navarro JM. Functional anatomy of thalamus and basal ganglia. Child Nerv Syst. 2002;18(8):386-404.
12. Aggleton JP. Multiple anatomical systems embedded within the primate medial temporal lobe: Implications for hippocampal function. Neurosci Biobehav R. 2012;36(7):1579-96.
13. Carrera E, Bogousslavsky J. The thalamus and behavior: effects of anatomically distinct strokes. Neurology. 2006;66(12):1817-23.
14. Mitchell AS, Sherman SM, Sommer MA, Mair RG, Vertes RP, Chudasama Y. Advances in understanding mechanisms of thalamic relays in cognition and behavior. J Neurosci. 2014;34(46):15340-6.
Figures