3703
Network structure of the mouse brain connectome with voxel resolution1Center for Mind/Brain Sciences, University of Trento, Rovereto, Italy, 2Functional Neuroimaging Laboratory, Center for Neuroscience and Cognitive Systems @ UniTn, Istituto Italiano di Tecnologia (IIT), Rovereto, Italy, 3Multimodal Imaging and Connectome Analysis Lab, Montreal Neurological Institute, McGill University, Montreal, QC, Canada
Synopsis
We provide a fine-grained description of the network structure and topology of the mouse connectome with voxel-resolution. We show that structural connectivity hubs can be spatially segregated into a dichotomic set of neural sink and neural source regions, and that functional and structural hubs show concordant neuro-anatomical topography. We finally demonstrate that the modular structure of the mouse connectome recapitulates large-scale fMRI networks of the mouse brain, including a default mode network core community, which is characterized by high structural resilience. Our results lay the foundation for topologically-driven targeted manipulations of the mouse connectome.
Introduction
Studies examining the structural and functional architecture of the mammalian brain have revealed the presence of specialized sub-systems (modules) interlinked by a small number of highly-connected “hub nodes”1,2, a configuration that enables the processing of specialized information as well as the effective integration of neural signals across different domains. Detailed investigations of the mouse brain connectome3 have produced ample evidence that the mouse brain recapitulates global organizational principles similar to those observed in higher species4. Replicating human investigations, recent studies have also provided initial evidence of spatial correlation between structural and functional anatomy inferred with resting-state fMRI (rsfMRI)5,6. However, investigations of the mouse connectome have to date relied on macroscopic parcellations, limiting the spatial resolution of connectional mapping and imposing arbitrary boundaries between regions. Furthermore, a precise characterization of the network structure and directional topography of the mouse brain is lacking. The present work leveraged a novel data driven model of the mouse connectome7 to provide a first-of-its-kind description of the network structure and directional topology of the mouse connectome with voxel-resolution, and relate some of the identified functional topographies to large-scale rsfMRI networks of the mouse brain.Methods
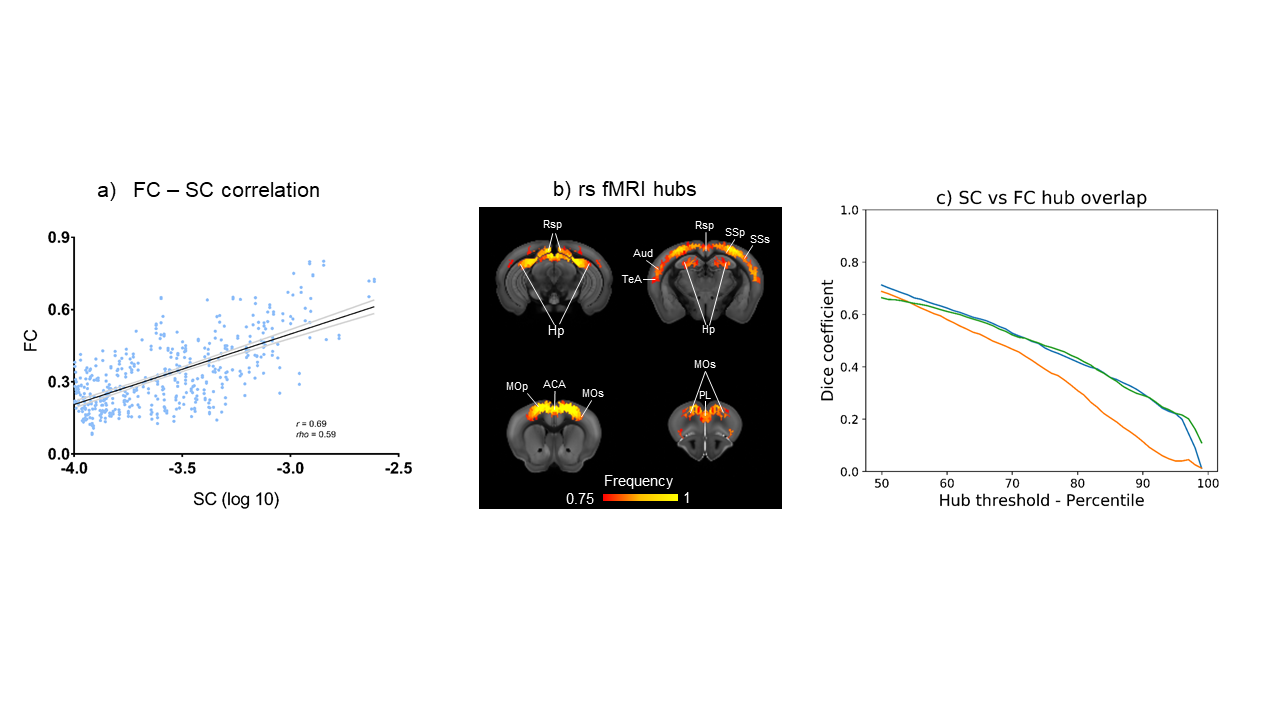
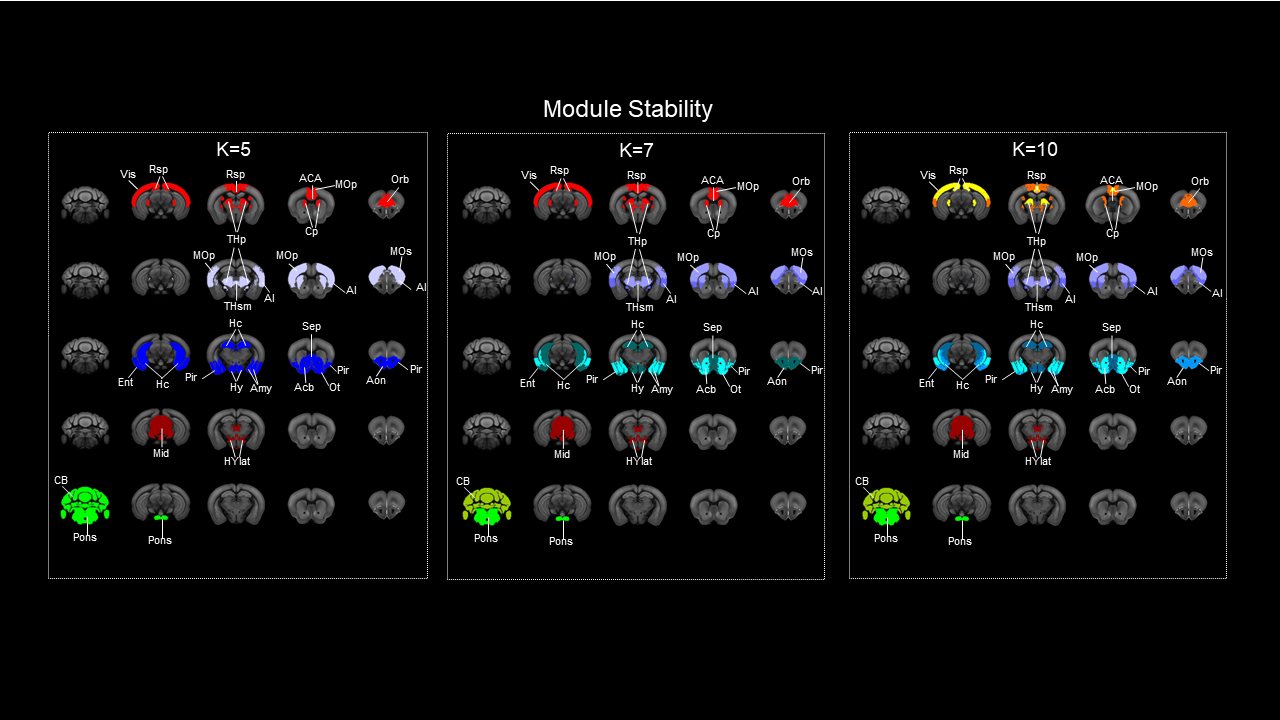
Generation of the voxel-wise mouse connectome. We estimated structural connectivity (SC) using a downsampled voxel scale model of the mouse structural connectome by aggregating neighbouring voxels according to a Voronoi diagram. This resulted in a weighted and directed 15’358x15’358 matrix that is computationally tractable, while retaining fine spatial features without the constraints of predefined anatomical boundaries. Percolation analysis was used to threshold weak links8. Graph theoretical analysis. We mapped out-strength (source), in-strength (sink), and global (source + sink) structural hubs. RsfMRI hubs were derived from 169 wild type mice2,9,10,11,12,13 using global connectivity, an aggregated index of functional connectivity9. For each measure, we iteratively computed the fraction of times a node scored among the top 50% to 99% of highest ranking nodes, and at every threshold used for hub identification, we quantified the spatial overlap between functional and structural hubs using the Dice coefficient. Structure function relationship was mapped by parcellating SC into a set of 74 aggregate-regions and using a non-directional square matrix. Module identification. Network modules were identified using the Louvain algorithm14. Modular detection was replicated using hierarchical clustering of a matrix obtained by computing between-nodes similarity based on the connectivity profile of each node.Results
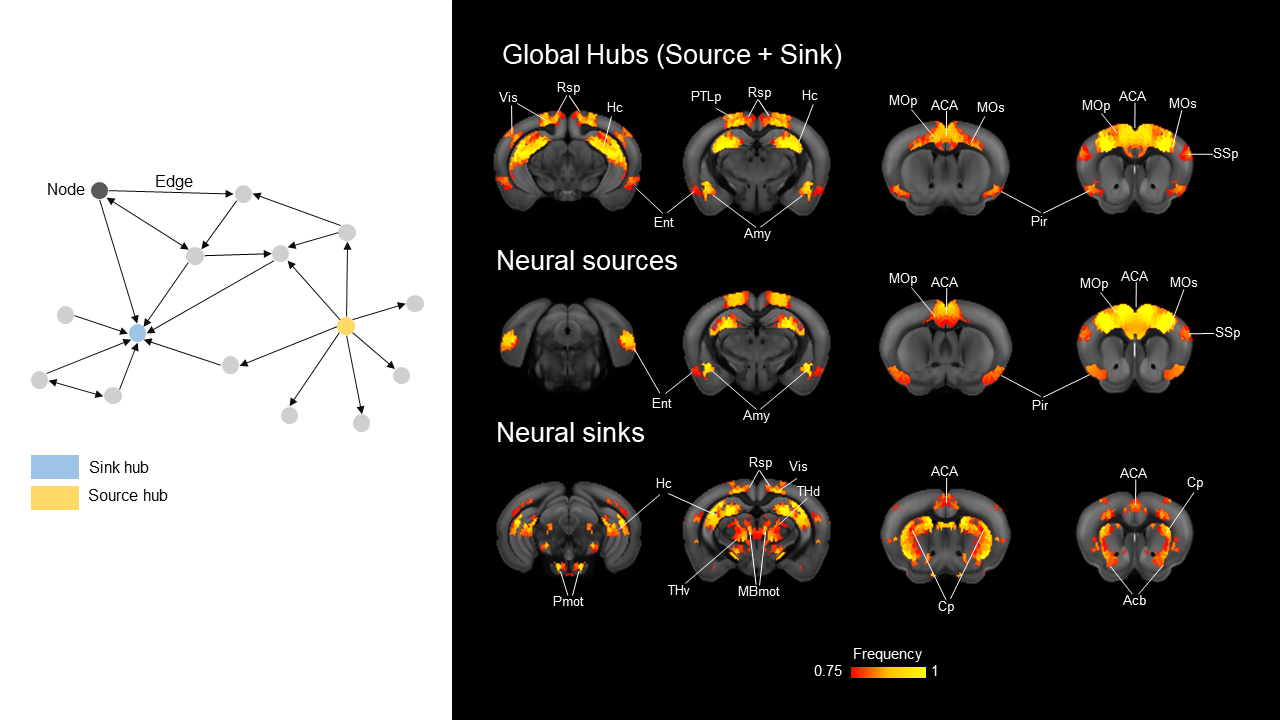
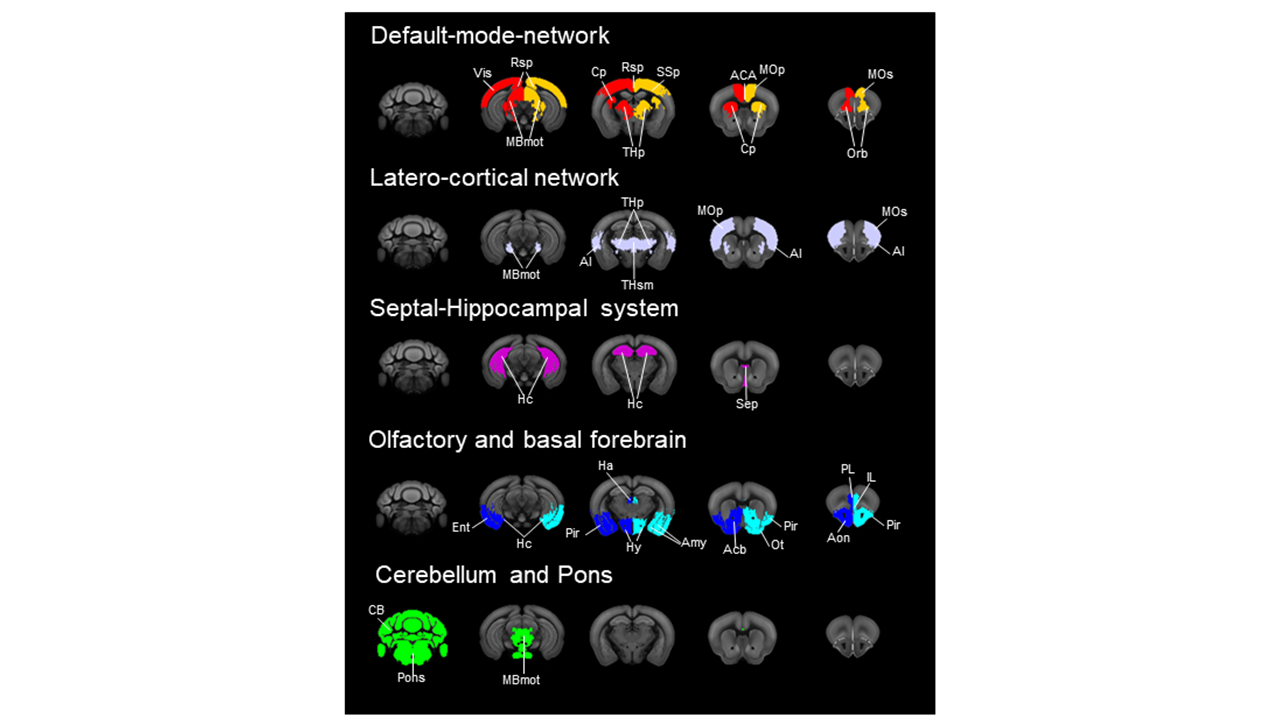
High connectivity strength hubs were predominantly located in the PFC, dorsal hippocampus, and olfactory-amygdaloyd areas (Fig 1). Neural sources were mainly located in the dorsal hippocampus and cortical regions of the mouse Default mode network (DMN, Fig. 1). Sink hubs (i.e., nodes with more ingoing than outgoing links) were mainly located in the thalamus, striatum, and ventral hippocampus (Fig. 1). A robust positive correlation was observed between whole rsfMRI and structural connectivity, and spatially concordant structural-functional topographies encompassed hub regions in fronto-cortical and hippocampal areas (Fig. 2a and 2b, respectively). Community detection using the modularity maximization approach resulted in seven large scale modules whose spatial topography closely recapitulates previously characterized rsfMRI networks of the mouse brain (Fig. 3). Hierarchical clustering replicated most of the prominent features observed with the modular detection, the only notable difference being the presence of a joint hippocampal basal forebrain module at low k (Fig 4). The DMN and midbrain networks maintained their structural integrity across a range of different solutions, whereas all the other network exhibited lower integrity and separated into smaller components with increasing k.Discussion and Conclusion
Directional hub mapping revealed a hierarchical organization of the mouse brain in which associative higher order cortical areas and olfactory regions serve as primary sources of neural input to the rest of the brain. By contrast, thalamo-cortical regions appear to be configured as neural sinks, and as such serve as major receivers of distributed neural input. The observation of connectional correlates of large scale rsfMRI networks corroborate the structural basis of rsfMRI coupling and suggest that long range coupling in functional macro-networks such as the DMN, or latero-cortical network is constrained by long-range projecting populations. Within this framework, we show that the mouse default mode network exhibits core-like properties, a feature consistent with this network as being critical for the integrity of brain function across different species, and serving as an entry-point for functional disconnectivity. By establishing a novel hierarchical description of the mouse brain connectome, our work advances our understanding of the network topology of the mammalian brain, and lays the foundation for target manipulation of the mouse connectome based on its topological properties.Acknowledgements
No acknowledgement found.References
1. van den Heuvel, M. P., & Sporns, O. (2013). Network hubs in the human brain. Trends in cognitive sciences, 17(12), 683-696.
2. Liska, A., Galbusera, A., Schwarz, A. J., & Gozzi, A. (2015). Functional connectivity hubs of the mouse brain. Neuroimage, 115, 281-291.
3. Oh, S. W., Harris, J. A., Ng, L., Winslow, B., Cain, N., Mihalas, S., ... & Mortrud, M. T. (2014). A mesoscale connectome of the mouse brain. Nature, 508(7495), 207.
4. Betzel, R. F., & Bassett, D. S. (2018). Specificity and robustness of long-distance connections in weighted, interareal connectomes. Proceedings of the National Academy of Sciences, 115(21), 4880-4889.
5. Stafford, J. M., Jarrett, B. R., Miranda-Dominguez, O., Mills, B. D., Cain, N., Mihalas, S., ... & Fryer, J. D. (2014). Large-scale topology and the default mode network in the mouse connectome. Proceedings of the National Academy of Sciences, 111(52), 18745-18750.
6. Grandjean, J., Zerbi, V., Balsters, J., Wenderoth, N., & Rudina, M. (2017). The structural basis of large-scale functional connectivity in the mouse. Journal of Neuroscience, 37(34), 8092-8101.
7. Knox, J. E., Harris, K. D., Graddis, N., Whitesell, J. D., Zeng, H., Harris, J. A., ... & Mihalas, S. (2018). High resolution data-driven model of the mouse connectome. bioRxiv.
8. Bordier, C., Nicolini, C., & Bifone, A. (2017). Graph analysis and modularity of brain functional connectivity networks: searching for the optimal threshold. Frontiers in neuroscience, 11, 1-9.
9. Liska, A., Bertero, A., Gomolka, R., Sabbioni, M., Galbusera, A., Barsotti, N., ... & Gozzi, A. (2017). Homozygous loss of autism-risk gene CNTNAP2 results in reduced local and long-range prefrontal functional connectivity. Cerebral cortex, 28(4), 1141-1153.
10. Suetterlin, P., Hurley, S., Mohan, C., Riegman, K. L., Pagani, M., Caruso, A., ... & Yee, Y. (2018). Altered neocortical gene expression, brain overgrowth and functional over-connectivity in Chd8 haploinsufficient mice. Cerebral Cortex, 28(6), 2192-2206.
11. Bertero, A., Liska, A., Pagani, M., Parolisi, R., Masferrer, M. E., Gritti, M., ... & Buffelli, M. (2018). Autism-associated 16p11.2 microdeletion impairs prefrontal functional connectivity in mouse and human. Brain, 141, 2055-2065
12. Pagani, M., Bertero, A., Liska, A., Galbusera, A., Sabbioni, M., Scattoni, M. L., ... & Gozzi, A. (2018). Deletion of autism risk gene Shank3 disrupts prefrontal connectivity. bioRxiv, 409284.
13. Michetti, C., Caruso, A., Pagani, M., Sabbioni, M., Medrihan, L., David, G., ... & Scattoni, M. L. (2017). The knockout of Synapsin II in mice impairs social behavior and functional connectivity generating an ASD-like phenotype. Cerebral Cortex, 27(10), 5014-5023.
14. Rubinov, M., & Sporns, O. (2010). Complex network measures of brain connectivity: uses and interpretations. Neuroimage, 52(3), 1059-1069.
Figures