3695
Sound-evoked functional magnetic resonance imaging of tinnitus and hyperacusis neural mechanisms1Department of Electrical and Electronic Engineering, The University of Hong Kong, Pokfulam, Hong Kong, 2Department of Communicative Disorders and Sciences, University at Buffalo, Buffalo, NY, United States, 3Department of Physics, City University of Hong Kong, Kowloon Tong, Hong Kong
Synopsis
Tinnitus involves the perception of sound in the absence of external acoustic stimuli. The underlying mechanisms of tinnitus have been investigated using noninvasive, whole brain functional imaging methods. However, the changes in the subcortical auditory system are less well understood. To address this knowledge gap, we conduct an auditory fMRI on a well-established sodium salicylate-induced rodent tinnitus and hyperacusis model. The results reveal frequency-dependent changes in auditory subcortical structures, inferior colliculus and lateral lemniscus by salicylate induced tinnitus. This study has advanced our understanding of subcortical auditory processing during tinnitus/hyperacusis and advances the rat model for further fMRI investigations.
Introduction
Tinnitus involves the perception of sound in the absence of external sound. Patients with tinnitus have considerable difficulties with concentration, productivity, and related activities. Tinnitus was traditionally thought to originate in the cochlear (1) and inner ear (2). Recently, neuroimaging studies observed aberrant neural activity within the central auditory pathway in tinnitus subjects (3-7). Functional imaging in tinnitus patients have further implicated the involvement of subcortical structures (6,8-9). However, how the subcortical auditory structures behave in tinnitus subjects largely remain unknown. To address the knowledge gap, here we utilize auditory fMRI to investigate the roles of subcortical auditory structures, inferior colliculus and lateral lemniscus, in the established sodium salicylate-induced rat tinnitus and hyperacusis model.Methods
Animal preparation (N=9): Sodium salicylate (SS) dissolved in saline solution (dosage: 250mg/kg; concentration: 50mg/ml) was intraperitoneally injected into adult SD rats. All subjects underwent auditory fMRI under light isoflurane (~1.0%), 2 hrs after saline and SS administration.
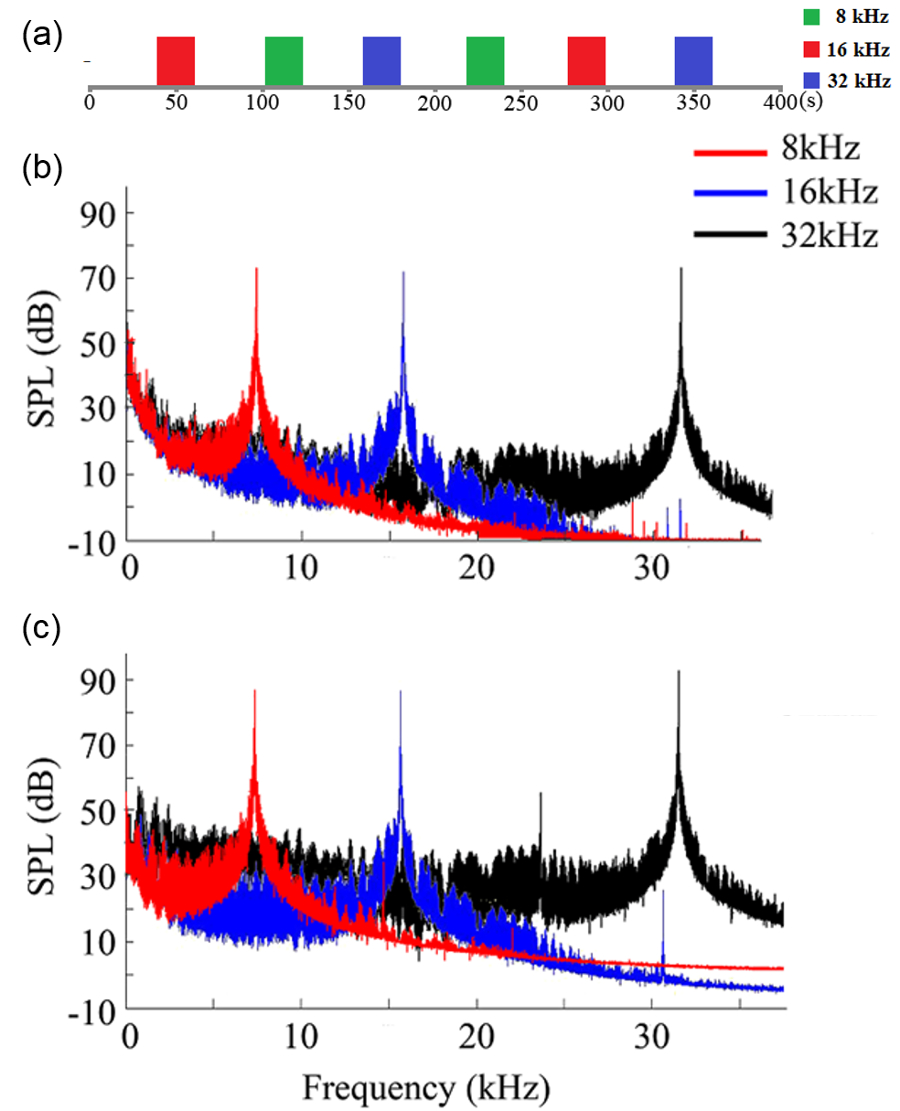
Auditory stimulation: Sound stimulation was delivered via a custom nylon tube inserted in the left ear and through a block-design paradigm (20s sound-on, 40s sound-off). Three frequencies (8, 16, 32khz) were presented in pseudorandom order for equal durations (Figure 1). Sound pressure level (SPL) 72 dB was used before salicylate administration and increased to 87, 87 and 92 dB (8, 16 and 32 kHz, respectively) after administration to compensate for the hearing threshold shift (10).
fMRI acquisition and analysis: fMRI data was acquired at 7T using GE-EPI (FOV=32×32mm2, matrix=64×64, α=56°, TE/TR=20/1000ms, 6 slices with 1.2mm thickness and 0.2mm gap). Standard fMRI preprocessing was performed before GLM analysis was applied to identify significant BOLD responses (p<0.001).
Results
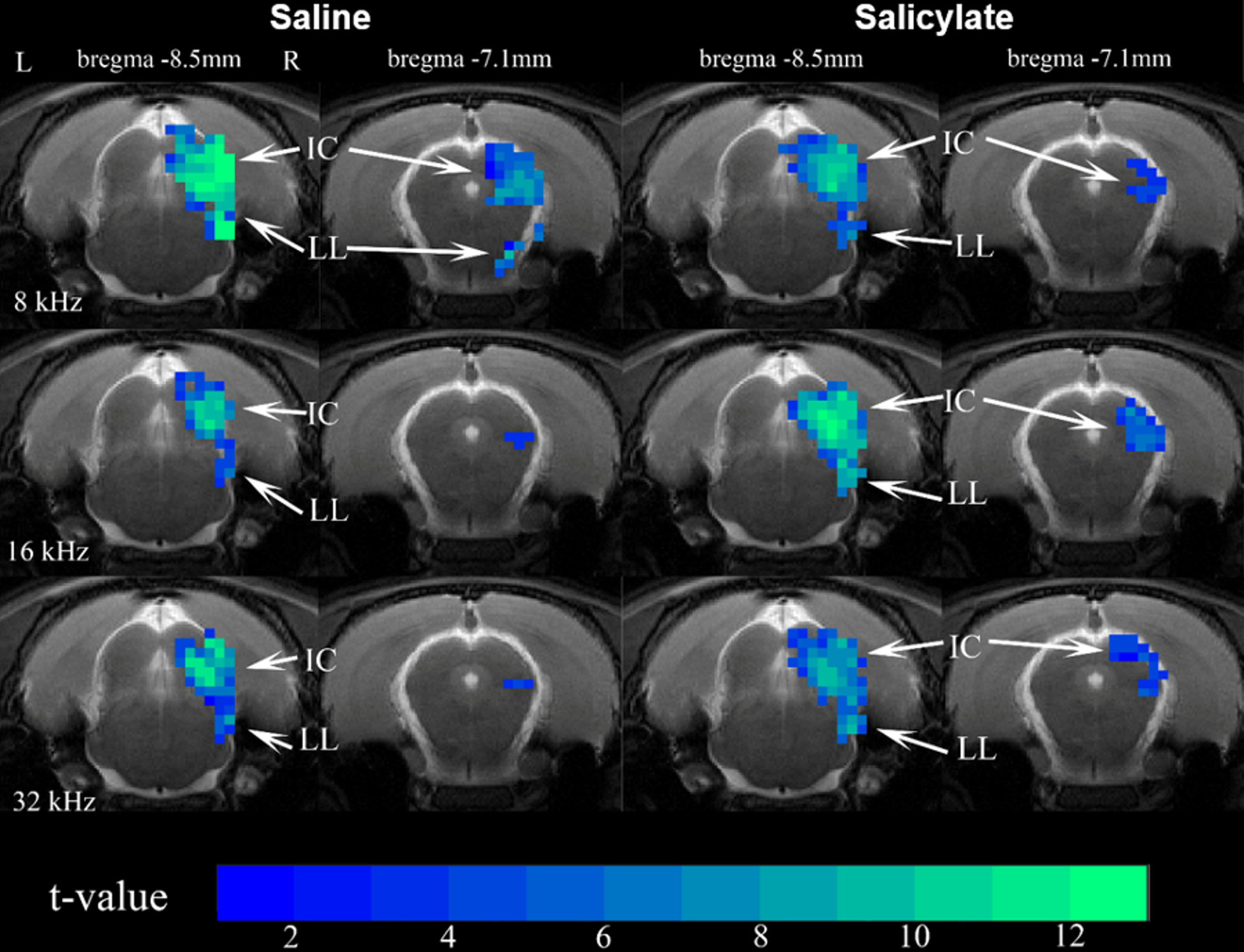
Figure 2 shows the sound-evoked BOLD maps before and after sodium salicylate (SS) injection. Positive BOLD responses are observed in IC and LL at all frequencies. In general, SS injection slightly reduces fMRI responses during 8kHz, slightly enhances the responses at 32kHz and leads to a great enhancement at 16kHz in IC and LL.
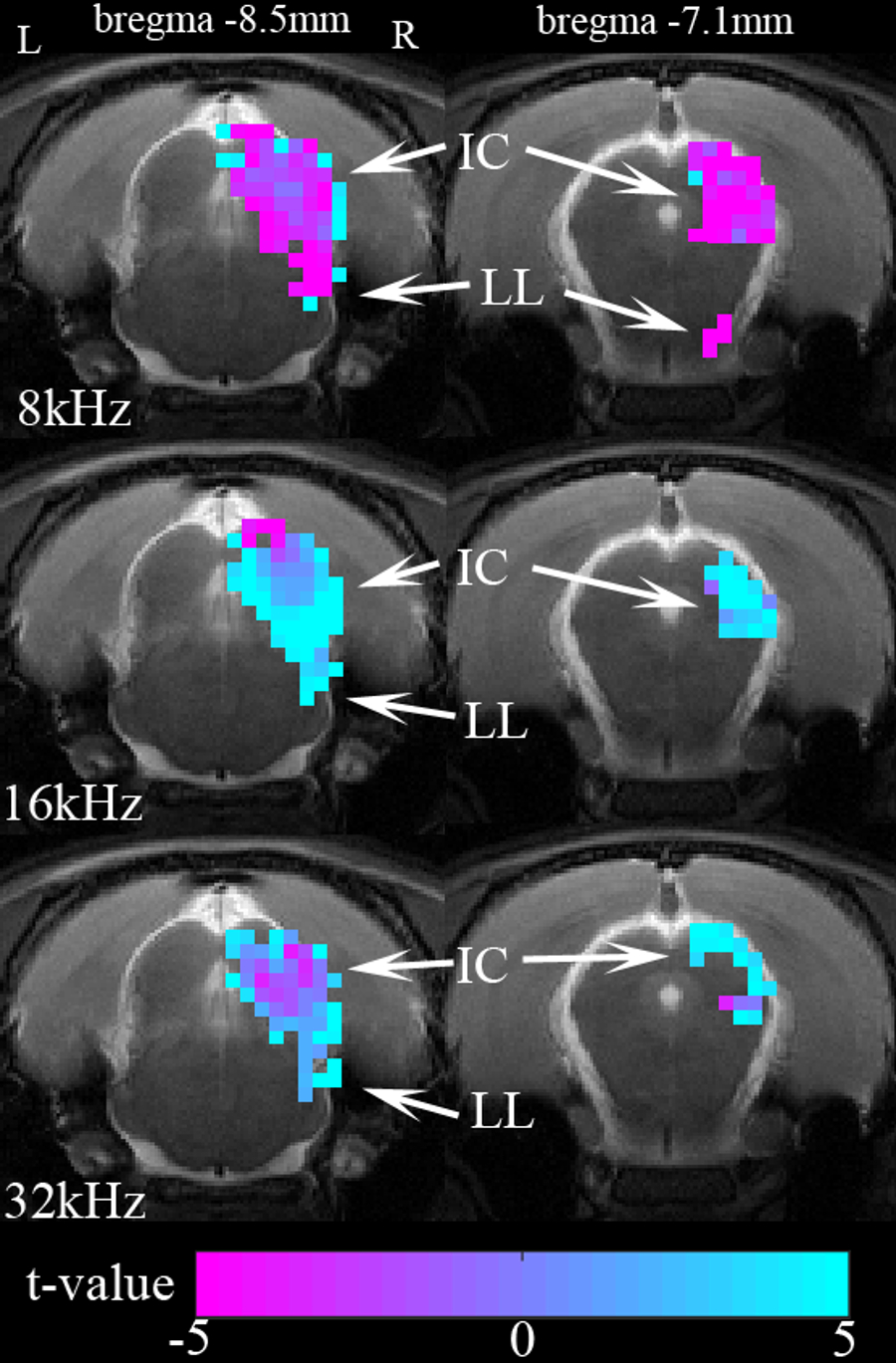
Figure 3 shows the averaged t-value difference maps before and after SS injection during 8, 16, and 32kHz. The differences were computed by subtracting the saline map from the salicylate map. In general, SS results in increased BOLD responses in the IC and LL during 16 and 32kHz and reduced responses during 8kHz.
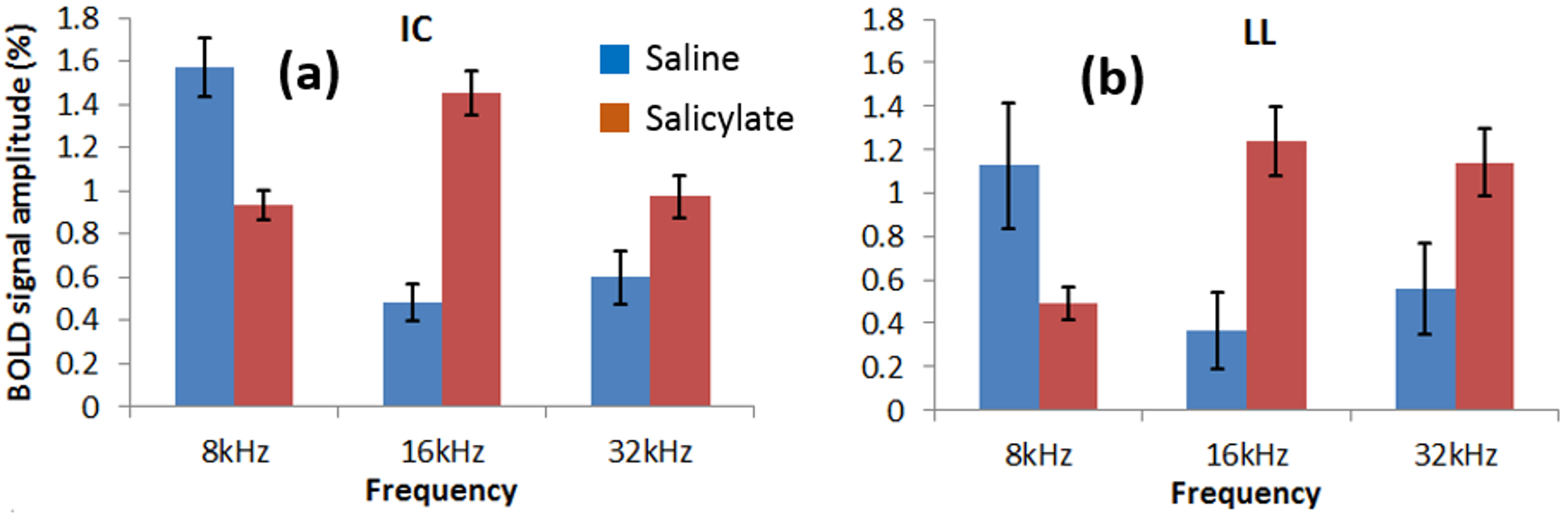
Figure 4 shows the BOLD signal amplitudes in the IC and LL before and after SS. The amplitudes are significantly increased at 16 (IC: p<0.001; LL: p<0.01) and 32kHz (IC: p<0.05; LL: p<0.01) and decreased at 8kHz (IC: p<0.05; LL: p<0.01) after SS. The largest difference observed in the IC is during 16kHz.
Discussion
This study demonstrates the roles of two major subcortical auditory structures, the inferior colliculus (IC) and lateral lemniscus (LL) (11-14), in tinnitus/hyperacusis induced by acute sodium salicylate (SS) administration. BOLD signal amplitudes increased significantly during 16 and 32kHz stimulation after SS and reduced slightly during 8kHz. The results suggest that SS induced tinnitus/hyperacusis leads to larger enhancements of subcortical responses at 16kHz, smaller enhancements at 32kHz, and reduced responses at 8kHz.
In our results, the BOLD signal amplitude difference is frequency dependent. The greatest increase in the IC after SS was during 16kHz, which had previously been behaviorally observed as approximately the salicylate-induced tinnitus pitch (15,16). Tonotopic reorganization in IC after salicylate administration may play a role for inducing such frequency dependent fMRI signal changes. Previous electrophysiological recordings in the rat auditory cortex observed tonotopic reorganization following SS. Neurons with characteristic frequency (CF) below 10kHz and above 20kHz shifted their CFs into the 10 to 20kHz range while the neurons with initial CFs between 10 and 20kHz showed minimal shift (17). Similar cortical tonotopic reorganization of tinnitus frequency had been observed in tinnitus patients (18). Furthermore, frequency receptive fields in the amygdala were found to be altered by local salicylate exposure. Neural activity was depressed at frequencies below 10kHz and above 20kHz, but greatly enhanced at frequency from 10 to 20kHz (19).
Conclusion
Our results showed the frequency dependent BOLD signal changes in IC and LL after salicylate administration, which likely extends the observation of subcortical tonotopic reorganization following salicylate administration. The findings align with and build upon previous behavioral and electrophysiological findings. The present study also demonstrated a fMRI protocol for rodent tinnitus/hyperacusis research.Acknowledgements
This research was supported by the Hong Kong General Research Fund (#21201217, #661313) and Hong Kong Health and Medical Research Fund (#11122581) grants to C Lau.References
[1] Kiang, N.Y., Moxon, E.C., Levine, R.A., 1970. Auditory-nerve activity in cats with normal and abnormal cochleas. In: Sensorineural hearing loss. Ciba Found Symp, 241-273.
[2] Jastreboff, P.J., 1990. Phantom auditory perception (tinnitus): Mechanisms of generation and perception. Neurosci Res 8, 221-254.
[3] Eggermont, J.J., Roberts, L.E., 2004. The neuroscience of tinnitus. Trends Neurosci 27, 676-682.
[4] Mirz, F., Pedersen, B., Ishizu, K., Johannsen, P., Ovesen, T., Stodkilde-Jorgensen, H., Gjedde, A., 1999. Positron emission tomography of cortical centers of tinnitus. Hear Res 134, 133-144.
[5] Norena, A.J., Eggermont, J.J., 2003. Changes in spontaneous neural activity immediately after an acoustic trauma: implications for neural correlates of tinnitus. Hear Res 183, 137-153.
[6] Roberts, L.E., Eggermont, J.J., Caspary, D.M., Shore, S.E., Melcher, J.R., Kaltenbach, J.A., 2010. Ringing ears: the neuroscience of tinnitus. J Neurosci 30, 14972-14979.
[7] Weisz, N., Moratti, S., Meinzer, M., Dohrmann, K., Elbert, T., 2005. Tinnitus perception and distress is related to abnormal spontaneous brain activity as measured by magnetoencephalography. PLoS Med 2, e153.
[8] Muhlau, M., Rauschecker, J.P., Oestreicher, E., Gaser, C., Rottinger, M., Wohlschlager, A.M., Simon, F., Etgen, T., Conrad, B., Sander, D., 2006. Structural brain changes in tinnitus. Cereb Cortex 16, 1283-1288.
[9] Boyen, K., de Kleine, E., van Dijk, P., & Langers, D. R. (2014). Tinnitus-related dissociation between cortical and subcortical neural activity in humans with mild to moderate sensorineural hearing loss. Hearing Research, 312, 48-59.
[10] Stolzberg, D., Chen, G.D., Allman, B.L., Salvi, R.J., 2011. Salicylate-induced peripheral auditory changes and tonotopic reorganization of auditory cortex. Neuroscience 180, 157-164.
[11] Cheung, M.M., Lau, C., Zhou, I.Y., Chan, K.C., Cheng, J.S., Zhang, J.W., Ho, L.C., Wu, E.X., 2012a. BOLD fMRI investigation of the rat auditory pathway and tonotopic organization. Neuroimage 60, 1205-1211.
[12] Lau, C., Pienkowski, M., Zhang, J.W., McPherson, B., Wu, E.X., 2015a. Chronic exposure to broadband noise at moderate sound pressure levels spatially shifts tone-evoked responses in the rat auditory midbrain. Neuroimage 122, 44-51.
[13] Lau, C., Zhang, J.W., McPherson, B., Pienkowski, M., Wu, E.X., 2015b. Long-term, passive exposure to non-traumatic acoustic noise induces neural adaptation in the adult rat medial geniculate body and auditory cortex. Neuroimage 107, 1-9.
[14] Zhang, J.W., Lau, C., Cheng, J.S., Xing, K.K., Zhou, I.Y., Cheung, M.M., Wu, E.X., 2013. Functional magnetic resonance imaging of sound pressure level encoding in the rat central auditory system. Neuroimage 65, 119-126.
[15] Kizawa, K., Kitahara, T., Horii, A., Maekawa, C., Kuramasu, T., Kawashima, T., Nishiike, S., Doi, K., Inohara, H., 2010. Behavioral assessment and identification of a molecular marker in a salicylate-induced tinnitus in rats. Neuroscience 165, 1323-1332.
[16] Yang, G., Lobarinas, E., Zhang, L., Turner, J., Stolzberg, D., Salvi, R., Sun, W., 2007. Salicylate induced tinnitus: behavioral measures and neural activity in auditory cortex of awake rats. Hear Res 226, 244-253.
[17] Chen, G.D., Stolzberg, D., Lobarinas, E., Sun, W., Ding, D., Salvi, R., 2013. Salicylate-induced cochlear impairments, cortical hyperactivity and re-tuning, and tinnitus. Hear Res 295, 100-113.
[18] Muhlnickel, W., Elbert, T., Taub, E., Flor, H., 1998. Reorganization of auditory cortex in tinnitus. Proc Natl Acad Sci U S A 95, 10340-10343.
[19] Chen, G.D., Manohar, S., Salvi, R., 2012. Amygdala hyperactivity and tonotopic shift after salicylate exposure. Brain Res 1485, 63-76.
Figures