3694
Tuning Neuromodulation Effects by Orientation Selective Deep Brain Stimulation in the Rat Medial Frontal Cortex1Center Magnetic Resonance Research, University of Minnesota, Minneapolis, MN, United States, 2First Department of Neurology, Faculty of Medicine, Masaryk University and St. Anne’s Teaching Hospital, Brno, Czech Republic, 3A. I. Virtanen Institute for Molecular Sciences, University of Eastern Finland, Kuopio, Finland, 4Department of Biomedical Engineering, University of Minnesota, Minneapolis, MN, United States, 5Division of Biostatistics, University of Minnesota, Minneapolis, MN, United States, 6Department of Neurosurgery, University of Minnesota, Minneapolis, MN, United States
Synopsis
With simultaneous fMRI and Deep Brain Stimulation (DBS), we demonstrate modulation of activity in rat amygdala by using axon Orientation Selective Stimulation (OSS) DBS in rat infralimbic cortex, a homologue brain region for treating depression in humans. Our results demonstrate orientation selectivity based on number of activated pixels especially in the amygdala, though qualitatively also other brain areas showed modulation based on stimulation angle. OSS may add a new level of optimization for treating major depression disorders.
Introduction
Thus far treating major depressive disorder using deep brain stimulation (DBS) has led to mixed results 1-3. This calls for better understanding of the targeted neural circuitry and inter-individual variability in clinical and cognitive phenotypes 4, which may be directly relevant for optimizing the stimulation parameters and the implantation site 5. Advances in electrode design 6 and stimulation paradigms 7,8 are promising avenues for optimizing stimulation outcomes. One such method is the recently introduced orientation selective stimulation (OSS) 9 that sensitizes stimulation to axons with different orientations. OSS was shown to be effective in the highly organized area of corpus callosum, but its effectiveness in more complex circuitry has not been investigated. The rat infralimbic cortex (IL) is the homologue of human subgenual cingulate cortex, a DBS target for treating major depression.10-14 IL sends strong connections to e.g. the medial frontal cortex, insula, medial thalamus, basal forebrain, amygdala and hypothalamus. Conventional electric stimulation would likely touch all these connections and thus influence all related networks.15 The objective of this study was to compare the network-level responses to IL DBS across multiple electric field orientations employing OSS and across multiple DBS frequencies using simultaneous fMRI.Methods
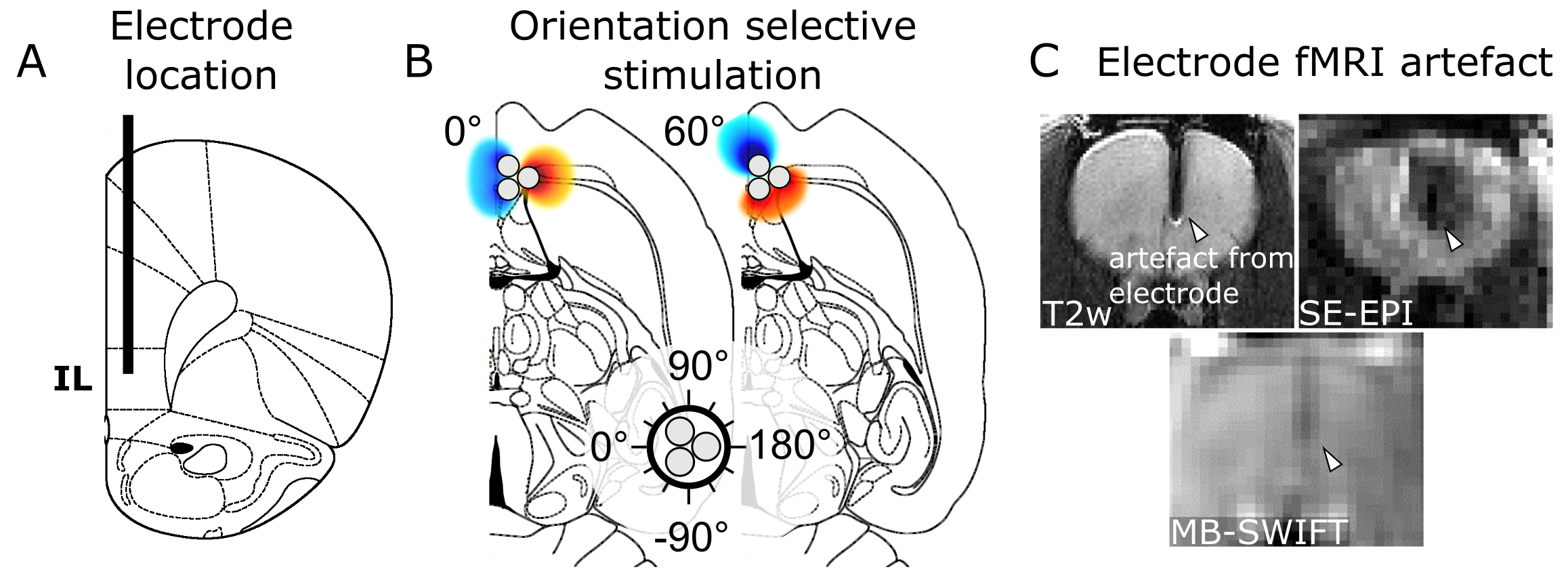
Three-channel tungsten wire electrodes (127 µm diameter per wire) were implanted in the rat IL (Fig 1A). For OSS, the stimulation angle was adjusted in 30° steps for a total of 12 fMRI acquisitions (Fig 1B; n = 8). The stimulation frequency dependence of the IL was tested with 20, 35, 70, 100, 130, 160 and 200 Hz (n = 6), and OSS was conducted using 20 Hz stimulation frequency. The waveform consisted of square, biphasic charge balanced pulses with no interpulse delay, 180 µs duration with amplitudes of 1.4 – 1.7 mA.
The fMRI stimulation paradigm consisted of 60 s of rest and 18 s of stimulation, repeated three times and ending in rest. Imaging was conducted at 9.4 T using Multi-Band SWeep Imaging with Fourier Transformation (MB-SWIFT; Fig 1C) 16,17 with the following parameters: TR = 0.97 ms, 3094 spokes per volume, resulting in temporal resolution of 3 s, bandwidth = 192 kHz, matrix size = 643, FOV = 3.5 x 3.5 x 6.4 cm3 and flip angle = 6°. Data analysis was performed using MATLAB, SPM5 and Aedes. Number of activated pixels and fMRI amplitude averaged over the animals were quantified using regions of interest (ROI).
Results
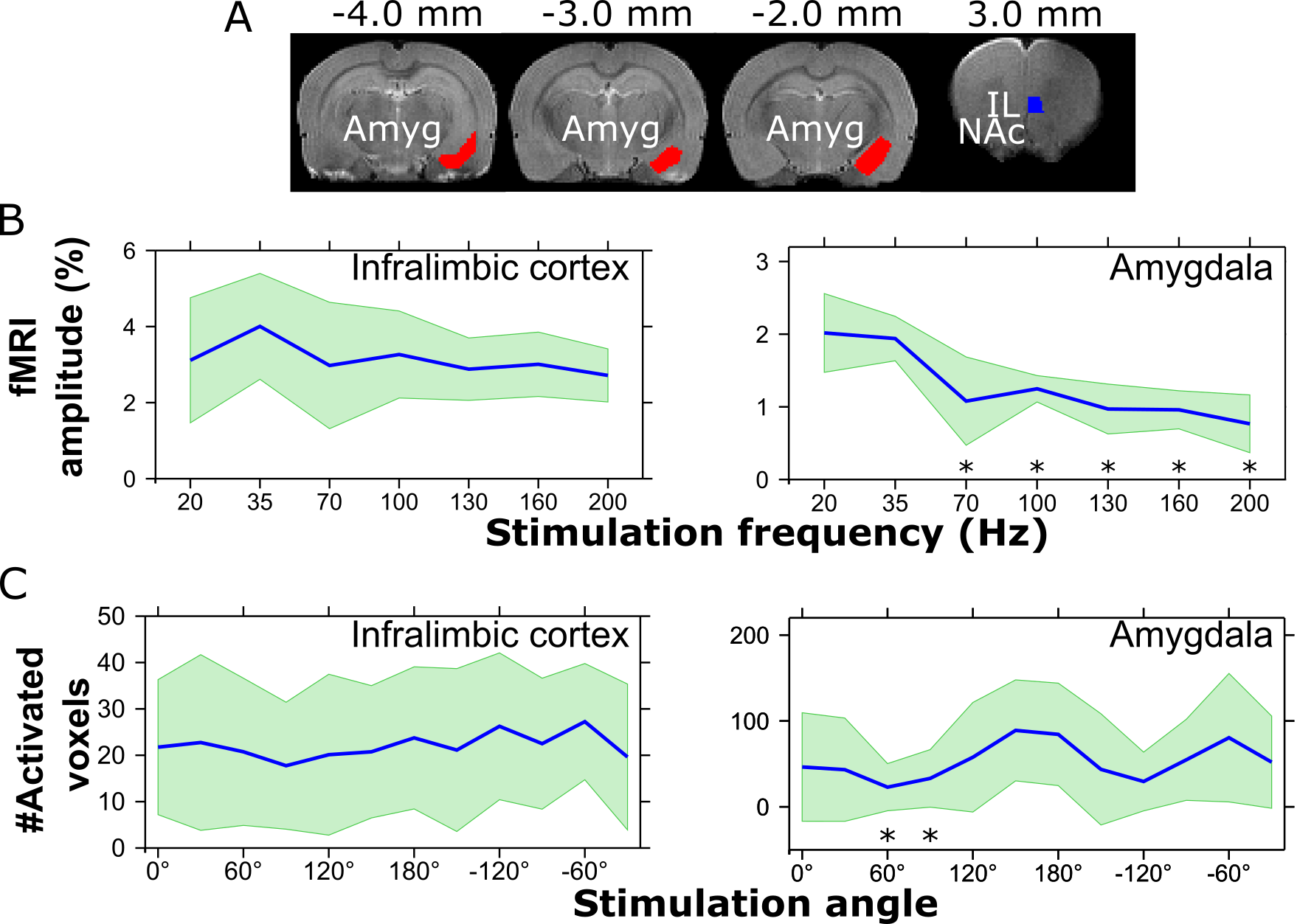
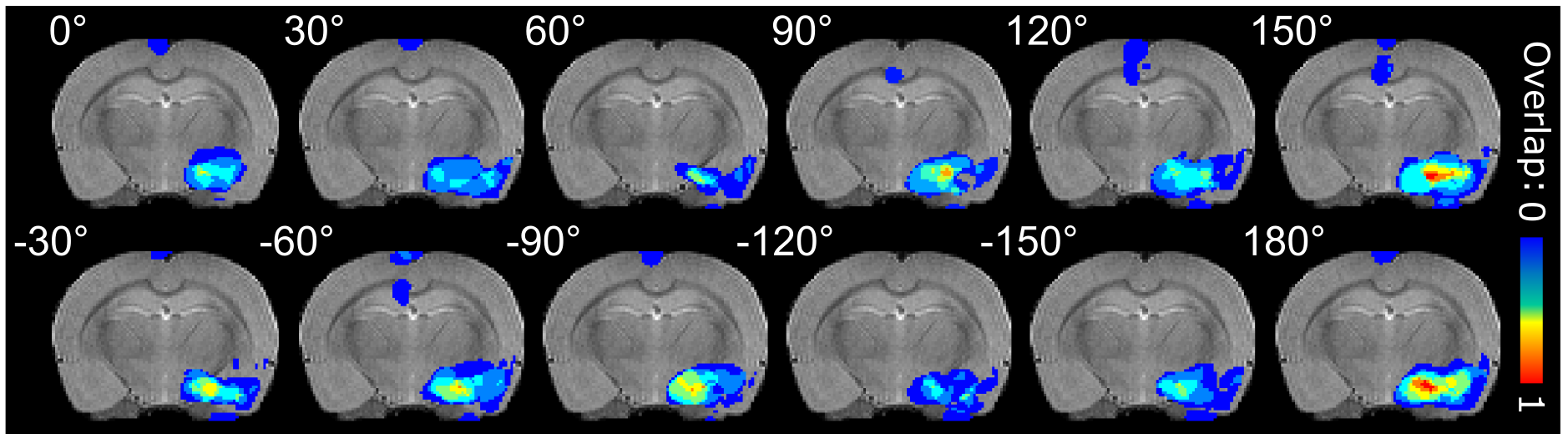
The stimulation activated widespread networks or brain regions known to be connected with the IL including e.g. the prefrontal cortex, medial orbitofrontal cortex, anterior cingulate cortex, anterior insula, olfactory tubercle and piriform cortex, basal forebrain structures, lateral septum and amygdala. ROI analyses were performed on IL and amygdala (Fig 2A). fMRI amplitude was seen to reduce with increasing stimulation frequency (Fig 2B), while the number of activated pixels did not show statistically significant differences with increasing frequency. On the other hand, when applying OSS, statistically significant orientation dependence was detected in the amygdala in number of activated pixels (Fig 2C), but not in fMRI amplitude. Overlapping individual animal activation maps 18 clearly show orientation dependence of stimulation in the amygdala (Fig 3). Qualitatively, modulation of activity based on the stimulation angle was also observed in various other brain regions.Discussion and Conclusion
The complexity of MDD DBS targets necessitates highly discriminatory modulation of desired pathways while ideally evading various high-risk neural circuits. DBS in IL has been shown effective even after destroying neuronal bodies and sparing axons using ibotenic acid in a rat model.12 This major role of axons in the DBS effect in IL indicates great potential for OSS paradigms capable of angle-dependent axonal stimulation.9 Indeed, our current findings show adjustable activation of the rat amygdala when applying OSS to the IL when looking at the number of activated pixels. The lack of effect in fMRI amplitude is likely related to the rather coarse electrode design where the large electrode stimulated a significant amount of the IL regardless of the stimulation angle. This may be alleviated with high-density silicon shanks.6
As the activation cluster near the electrode was insensitive to the changes of the stimulation angle but the terminal zone structures were differentially stimulated, our findings substantiate the ability of OSS to recruit neuronal pathways of distinct orientations relative to the position of the electrode, even in complex circuits such as those involved in MDD. OSS DBS technique may offer a new avenue for stimulus parameter optimization during DBS, so that relevant pathways are stimulated, while simultaneously avoiding crossing pathways associated with side effects.
Acknowledgements
This work was supported by the National Institutes of Health U01-NS103569-01, the Center for Magnetic Resonance Research NIH core grant P41-EB015894, P30 NS076408, NIH R01-NS094206, NIH R01-NS094206, WM KECK Foundation, the EU H2020 Marie Skłodowska RISE project #691110 (MICROBRADAM), Erkko foundation (OG), Academy of Finland (AS) and The Emil Aaltonen Foundation (LJL).References
1. Bergfeld, I. O., Mantione, M. et al. Deep brain stimulation of the ventral anterior limb of the internal capsule for treatment-resistant depression. JAMA Psychiatry 73, 456-464, doi:10.1001/jamapsychiatry.2016.0152 (2016).
2. Dougherty, D. D., Rezai, A. R. et al. A Randomized Sham-Controlled Trial of Deep Brain Stimulation of the Ventral Capsule/Ventral Striatum for Chronic Treatment-Resistant Depression. Biological Psychiatry 78, 240-248, doi:10.1016/j.biopsych.2014.11.023 (2015).
3. Youngerman, B. E. & Sheth, S. A. Deep brain stimulation for treatment-resistant depression: optimizing interventions while preserving valid trial design. Annals of Translational Medicine 5, S1, doi:10.21037/atm.2017.03.40 (2017).
4. Widge, A. S., Deckersbach, T. et al. Deep brain stimulation for treatment-resistant psychiatric illnesses: What has gone wrong and what should we do next? Biological Psychiatry 79, e9-e10, doi:10.1016/j.biopsych.2015.06.005 (2016).
5. Morishita, T., Fayad, S. M. et al. Deep Brain Stimulation for Treatment-resistant Depression: Systematic Review of Clinical Outcomes. Neurotherapeutics 11, 475-484, doi:10.1007/s13311-014-0282-1 (2014).
6. Tsai, D., John, E. et al. 7510-7513 (IEEE).
7. Martens, H. C. F., Toader, E. et al. Spatial steering of deep brain stimulation volumes using a novel lead design. Clinical neurophysiology 122, 558-566 (2011).
8. Chaturvedi, A., Foutz, T. J. et al. Current steering to activate targeted neural pathways during deep brain stimulation of the subthalamic region. Brain Stimulation: Basic, Translational, and Clinical Research in Neuromodulation 5, 369-377 (2012).
9. Lehto, L. J., Slopsema, J. P. et al. Orientation selective deep brain stimulation. Journal of neural engineering 14, 016016 (2017).
10. Diorio, D., Viau, V. et al. The role of the medial prefrontal cortex (cingulate gyrus) in the regulation of hypothalamic-pituitary-adrenal responses to stress. Journal of Neuroscience 13, 3839-3847 (1993).
11. Gabbott, P. L. A., Warner, T. A. et al. Areal and synaptic interconnectivity of prelimbic (area 32), infralimbic (area 25) and insular cortices in the rat. Brain research 993, 59-71 (2003).
12. Hamani, C., Diwan, M. et al. Antidepressant-Like Effects of Medial Prefrontal Cortex Deep Brain Stimulation in Rats. Biological Psychiatry 67, 117-124, doi:10.1016/j.biopsych.2009.08.025 (2010).
13. Ostrander, M. M., Richtand, N. M. et al. Stress and amphetamine induce Fos expression in medial prefrontal cortex neurons containing glucocorticoid receptors. Brain research 990, 209-214 (2003).
14. Takagishi, M. & Chiba, T. Efferent projections of the infralimbic (area 25) region of the medial prefrontal cortex in the rat: an anterograde tracer PHA-L study. Brain research 566, 26-39 (1991).
15. Hamani, C., Mayberg, H. et al. The subcallosal cingulate gyrus in the context of major depression. Biol Psychiatry 69, 301-308, doi:10.1016/j.biopsych.2010.09.034 (2011).
16. Idiyatullin, D., Corum, C. A. et al. Multi-Band-SWIFT. J Magn Reson 251, 19-25, doi:10.1016/j.jmr.2014.11.014 (2015).
17. Lehto, L. J., Idiyatullin, D. et al. MB-SWIFT functional MRI during deep brain stimulation in rats. NeuroImage 159, 443-448, doi:10.1016/j.neuroimage.2017.08.012 (2017).
18. Spiridon, M., Fischl, B. et al. Location and spatial profile of category-specific regions in human extrastriate cortex. Hum Brain Mapp 27, 77-89, doi:10.1002/hbm.20169 (2006).
Figures